Avertissements:
Apprenons à connaître le Cambium. Après avoir lu cet article, vous apprendrez à connaître : 1. L’origine du cambium 2. Cambium fasciculaire et inter-fasciculaire 3. Durée 4. Les fonctions 5. Structure 6. Division cellulaire 7. Épaississement dans les paumes.
Origine du cambium :
Le squelette vasculaire primaire est construit par la maturation des cellules des brins de procambium pour former le xylème et le phloème. Chez les plantes qui ne possèdent pas de croissance secondaire, toutes les cellules des brins de procambium mûrissent et se développent en tissu vasculaire.
Dans la plante qui possède une croissance secondaire ultérieure, une partie du brin de procambium reste méristématique et donne naissance au cambium proprement dit. Dans les racines, la formation du cambium diffère de celle des tiges en raison de la disposition radiale des brins alternés de xylème et de phloème.
Avertissements:
Ici, le cambium naît sous forme de bandes discrètes de tissu dans les brins de procambium à l’intérieur des groupes de phloème primaire. Plus tard, les bandes de cambium, par leur extension latérale, sont réunies dans le péricycle en face des rayons du xylème primaire. La formation des tissus secondaires est la plus rapide sous les groupes de phloème, de sorte que le cambium, vu dans la coupe transversale des racines âgées, forme bientôt un cercle.
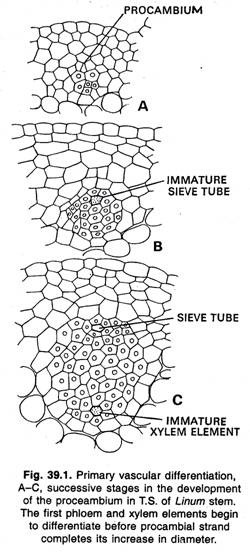
Cambium fasciculaire et interfasciculaire :
Dans les tiges, le premier procambium qui se développe à partir du promeristem se trouve généralement sous forme de brins isolés. Chez certaines plantes, ces brins primitifs deviennent rapidement, unis latéralement par des brins similaires supplémentaires formés entre eux et par l’extension latérale des brins primitifs.
Pendant la suite du développement, ce cylindre procambial donne naissance à un cylindre de tissu vasculaire primaire (xylème et phloème) et de cambium. Plus tard, un cylindre de tissu vasculaire secondaire se forme, qui naît en brins comme le cylindre primaire. Chez la renoncule et certaines autres plantes herbacées, les brins de procambium, et les tissus vasculaires primaires, ne fusionnent pas latéralement mais restent sous forme de brins distincts.
Avertissements:
Plus souvent dans les tiges herbacées, le cambium s’étend latéralement à travers les espaces intermédiaires jusqu’à ce qu’un cylindre complet soit formé. Lorsqu’une telle extension se produit, le cambium naît de cellules méristématiques inter-fasciculaires dérivées du méristème apical.
Les bandes de cambium qui naissent à l’intérieur des faisceaux collatéraux sont appelées cambium fasciculaire, et les bandes de cambium que l’on trouve entre les faisceaux sont appelées cambium inter-fasciculaire.
Durée du cambium :
La durée de la vie fonctionnelle du cambium varie beaucoup dans les différentes espèces et aussi dans les différentes parties d’une même plante. Chez une plante ligneuse pérenne, le cambium de la tige principale vit depuis sa formation jusqu’à la mort de la plante.
C’est seulement par l’activité continue du cambium dans la production de nouveaux xylème et phloème que ces plantes peuvent maintenir leur existence. Dans les feuilles, les inflorescences et autres parties caduques, la vie fonctionnelle du cambium est courte. Ici, toutes les cellules du cambium arrivent à maturité en tant que tissu vasculaire. Le xylème secondaire se trouve directement sur le phloème secondaire dans de tels faisceaux.
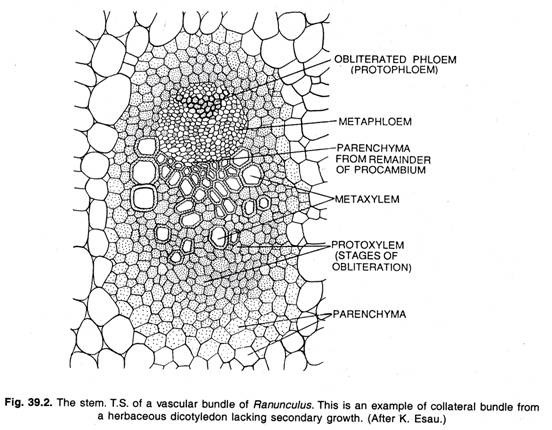
Fonction du cambium :
Le méristème qui forme les tissus secondaires est constitué d’une feuille unisériée de paraphes qui forment de nouvelles cellules généralement des deux côtés. Le cambium forme le xylème à l’intérieur et le phloème à l’extérieur. La division tangentielle de la cellule cambiale forme deux cellules filles apparemment identiques.
L’une des cellules filles reste méristématique, c’est-à-dire la cellule cambiale persistante, l’autre devient une cellule mère de xylème ou une cellule mère de phloème selon sa position interne ou externe à l’initiale. La cellule de cambium se divise continuellement de la même manière ; une cellule fille reste toujours méristématique, la cellule de cambium, tandis que l’autre devient soit une cellule mère de xylème, soit une cellule mère de phloème.
Probablement, il n’y a pas d’alternance définie et pendant de brèves périodes, un seul type de tissu est formé. Les cellules adjacentes du cambium se divisent presque en même temps, et les cellules filles appartiennent au même tissu. De cette façon, la continuité tangentielle du cambium est maintenue.
Structure du cambium :
Avertissements :
Il existe deux conceptions générales du cambium comme couche initiatrice :
1. Qu’il consiste en une couche uniseriate d’initiales permanentes avec des dérivés qui peuvent se diviser quelques fois et se transformer rapidement en tissu permanent ;
2. Qu’il y a plusieurs rangées de cellules initiatrices qui forment une zone de cambium, dont quelques rangées individuelles persistent comme couches cellulaires pendant un certain temps. Pendant les périodes de croissance, les cellules mûrissent continuellement des deux côtés du cambium, il devient tout à fait évident que seule une seule couche de cellules peut avoir une existence permanente en tant que cambium.
Les autres couches, si elles sont présentes, ne fonctionnent que temporairement et se transforment complètement en cellules permanentes. Au sens strict, seules les initiales constituent le cambium, mais fréquemment le terme est utilisé en référence à la zone cambiale, car il est difficile de distinguer les initiales de leurs dérivés récents.

Structure cellulaire du cambium :
Il existe deux types différents de cellules du cambium :
1. Les initiales de rayons, qui sont plus ou moins isodiamétriques et donnent naissance aux rayons vasculaires ; et
2. Les initiales fusiformes, les cellules effilées allongées qui se divisent pour former toutes les cellules du système vertical.
ADVERTISSEMENTS:
Les cellules cambiales sont fortement vacuolisées, généralement avec une grande vacuole et un cytoplasme périphérique mince. Le noyau est grand et dans les cellules fusiformes est très allongé. Les parois des cellules cambiales présentent des champs de puits primaires avec des plasmodesmes. Les parois radiales sont plus épaisses que les parois tangentielles, et leurs champs de fosses primaires sont profondément déprimés.

Division cellulaire dans le cambium :
Avec le résultat des divisions tangentielles (périclinales) des cellules du cambium, le phloème et le xylème sont formés. Les tissus vasculaires sont formés dans deux directions opposées, les cellules du xylème vers l’intérieur de l’axe, les cellules du phloème vers sa périphérie. Les divisions tangentielles des initiales du cambium pendant la formation des tissus vasculaires déterminent la disposition des dérivés du cambium en rangées radiales.
Avertissements:
Puisque la division est tangentielle, les cellules filles qui persistent en tant qu’initiales du cambium augmentent uniquement en diamètre radial. Les nouvelles initiales du cambium formées par des divisions transversales augmentent considérablement en longueur ; celles formées par des divisions radiales n’augmentent pas en longueur.
Alors que le cylindre du xylème augmente en épaisseur par croissance secondaire, le cylindre cambial augmente également en circonférence. La cause principale de cette croissance est l’augmentation du nombre de cellules dans la direction tangentielle, suivie d’une expansion tangentielle de ces cellules.
Croissance cambiale autour des plaies :
L’une des fonctions importantes du cambium est la formation de cals ou de tissus de blessures, et la guérison des blessures. Lorsque des blessures se produisent sur les plantes, une grande quantité de tissu parenchymateux mou se forme sur ou sous la surface blessée ; ce tissu est connu sous le nom de cal. Le cal se développe à partir du cambium et par la division des cellules du parenchyme dans le phloème et le cortex.
Durant le processus de guérison d’une blessure, le cal se forme. Dans celui-ci, il y a d’abord une prolifération abondante des cellules du cambium, avec la production d’un parenchyme massif. Les cellules externes de ce tissu se subérisent, ou le périderme se développe en leur sein, avec pour résultat la formation d’une écorce.
Cependant, juste sous cette écorce, le cambium reste actif et forme un nouveau tissu vasculaire de manière normale. Le nouveau tissu formé de manière normale étend la couche de croissance sur la blessure jusqu’à ce que les deux côtés opposés se rencontrent. Les couches de cambium s’unissent alors et la plaie devient complètement couverte.
Avertissements:
Cambium dans le bourgeonnement et la greffe :
Dans les pratiques de bourgeonnement et de greffage, le cambium de la souche et du scion donne lieu à un cal qui s’unit et développe une couche continue de cambium qui donne lieu à un tissu conducteur normal. Il y a une union réelle du cambium du stock et du scion de deux plantes pendant les pratiques de bourgeonnement et de greffage et donc ces pratiques ne sont pas couramment rencontrées chez les monocotylédones.
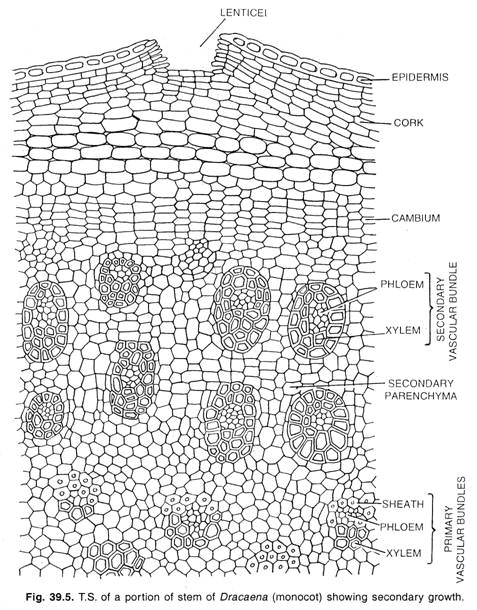
Cambium chez les monocotylédones :
Un type spécial de croissance secondaire se produit dans quelques formes monocotylédones, telles que Dracaena, Aloe, Yucca, Veratrum et quelques autres genres. Chez ces plantes, la tige augmente de diamètre en formant un cylindre de nouveaux faisceaux noyés dans un tissu.
Ici, une couche de cambium se développe à partir du parenchyme méristématique du péri-cycle ou des cellules les plus internes du cortex. Dans le cas des racines, le cambium de celle-ci se développe dans l’endoderme. Les initiales du cambium s’enroulent en gradins pour former un cambium étagé comme on le trouve dans le cambium normal de certaines dicotylédones.
Cambium en épaississement chez les palmiers :
Avertissements:
Les tiges des palmiers n’augmentent pas en circonférence, à cause d’une quelconque activité cambiale mais cet épaississement est le résultat d’une augmentation progressive de la taille des cellules et des espaces intercellulaires et parfois de la prolifération des tissus fibreux. C’est le type de croissance primaire continue de longue durée.
Le processus est le suivant :
La plupart des monocotylédones n’ont pas de croissance secondaire, mais avec le résultat d’une croissance primaire intense et longtemps continue, ils peuvent produire des corps aussi grands que ceux des palmiers. Les monocotylédones produisent souvent un épaississement rapide sous le méristème apical au moyen d’un méristème d’épaississement primaire périphérique comme le montre la figure.
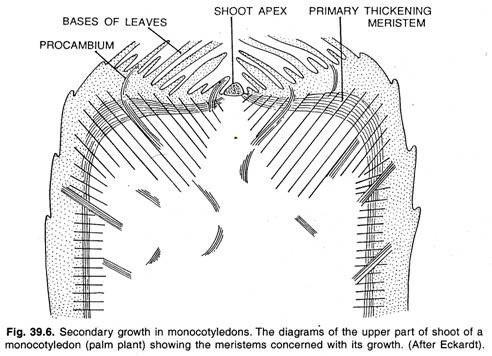
L’activité du méristème d’épaississement primaire ressemble à la croissance secondaire que l’on trouve chez certaines monocotylédones comme Dracaena, Yucca, etc. Le méristème apical également connu sous le nom d’apex des pousses ne produit qu’une petite partie du corps primaire, c’est-à-dire une colonne centrale de parenchyme et de brins vasculaires.
La majeure partie du corps de la plante est formée par le méristème primaire d’épaississement. Le méristème primaire d’épaississement se trouve sous la feuille-primordia, qui se divise de manière périclinale en produisant des rangées anticlinales de cellules. Ces cellules se différencient en un tissu formé de parenchyme terrestre traversé par des brins procambiaux.
Ces brins procambiaux se développent plus tard en faisceaux vasculaires. Les cellules du parenchyme terrestre s’agrandissent et se divisent de manière répétée, provoquant une augmentation de l’épaisseur. De cette façon, le méristème apical et le méristème primaire d’épaississement donnent naissance à la majeure partie des tissus de la tige des monocotylédones.
L’épaississement a lieu dans les monocotylédones, comme les palmiers, grâce aux activités du méristème apical et du méristème primaire d’épaississement.