ADVERTISEMENTS:
Aprendamos sobre el Cambium. Después de leer este artículo aprenderás sobre: 1. Origen del cámbium 2. Cambium fascicular e interfascicular 3. Duración 4. Funciones 5. Estructura 6. División celular 7. Engrosamiento en las palmas.
Origen del cambium:
El esqueleto vascular primario se construye mediante la maduración de las células de los filamentos del procambium para formar el xilema y el floema. Las plantas que no poseen crecimiento secundario, todas las células de los filamentos de procambium maduran y se convierten en tejido vascular.
En las plantas que tienen crecimiento secundario posterior, una parte de la hebra de procambium permanece meristemática y da lugar al cambium propiamente dicho. En las raíces la formación del cambium difiere de la de los tallos debido a la disposición radial de las hebras alternas de xilema y floema.
ADVERTENCIAS:
Aquí el cambium surge como tiras discretas de tejido en los filamentos de procambium dentro de los grupos de floema primario. Posteriormente, las tiras de cambium por su extensión lateral se unen en el periciclo frente a los rayos del xilema primario. La formación de tejido secundario es más rápida debajo de los grupos de floema, de modo que el cambium, como se ve en la sección transversal de las raíces más viejas, pronto forma un círculo.
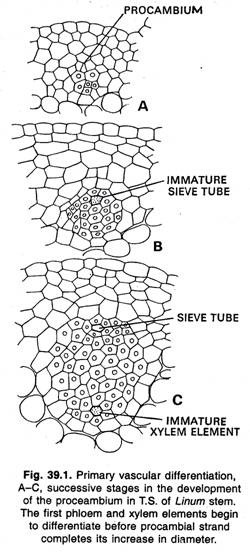
Cambium fascicular e interfascicular:
En los tallos el primer procambium que se desarrolla a partir del promeristema suele encontrarse en forma de filamentos aislados. En algunas plantas estos primeros filamentos formados pronto se unen lateralmente por filamentos similares adicionales formados entre ellos y por la extensión lateral de los primeros filamentos formados.
Durante el desarrollo posterior este cilindro procambial da lugar a un cilindro de tejido vascular primario (xilema y floema) y cambium. Más tarde se forma un cilindro de tejido vascular secundario que surge en hebras al igual que el cilindro primario. En Ranunculus y algunas otras plantas herbáceas, los filamentos de procambium, y los tejidos vasculares primarios, no se fusionan lateralmente sino que permanecen como filamentos discretos.
ADVERTENCIAS:
Más a menudo en los tallos herbáceos el cambium se extiende lateralmente a través de los espacios intermedios hasta que se forma un cilindro completo. Cuando se produce dicha extensión, el cambium surge de células meristemáticas interfasciculares derivadas del meristemo apical.
Las tiras de cambium que surgen dentro de los haces colaterales se conocen como cambium fascicular, y las tiras de cambium que se encuentran entre los haces se conocen como cambium interfascicular.
Duración del cambium:
La duración de la vida funcional del cambium varía mucho en diferentes especies y también en diferentes partes de la misma planta. En una planta leñosa perenne el cambium del tallo principal vive desde el momento de su formación hasta la muerte de la planta.
Sólo mediante la actividad continuada del cambium en la producción de nuevo xilema y floema pueden estas plantas mantener su existencia. En las hojas, inflorescencias y otras partes caducas, la vida funcional del cambium es corta. Aquí todas las células del cambium maduran como tejido vascular. El xilema secundario se encuentra directamente sobre el floema secundario en tales haces.
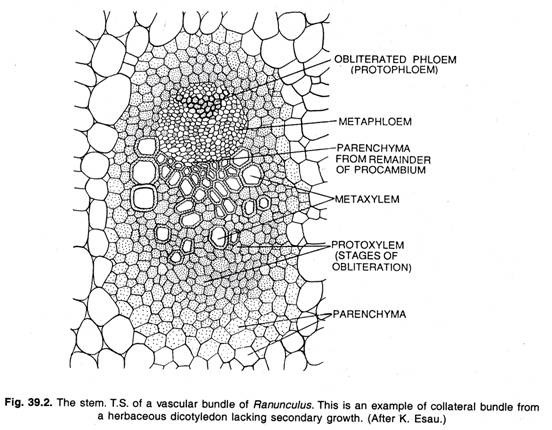
Función del cambium:
El meristemo que forma los tejidos secundarios consiste en una hoja uniseriada de iniciales que forman nuevas células generalmente en ambos lados. El cambium forma xilema internamente y floema externamente. La división tangencial de la célula cambial forma dos células hijas aparentemente idénticas.
Una de las células hijas permanece meristemática, es decir, la célula cambial persistente, la otra se convierte en célula madre del xilema o en célula madre del floema dependiendo de su posición interna o externa a la inicial. La célula cambial se divide continuamente de forma similar; una célula hija permanece siempre meristemática, la célula cambial, mientras que la otra se convierte en célula madre del xilema o del floema.
Probablemente no hay una alternancia definida y durante breves períodos sólo se forma un tipo de tejido. Las células adyacentes del cambium se dividen casi al mismo tiempo y las células hijas pertenecen al mismo tejido. De este modo, se mantiene la continuidad tangencial del cambium.
Estructura del cambium:
ADVERTENCIAS:
Hay dos concepciones generales del cambium como capa iniciadora:
1. Que consiste en una capa uniseriada de iniciales permanentes con derivados que pueden dividirse unas pocas veces y convertirse pronto en tejido permanente;
2. Que hay varias filas de células iniciadoras que forman una zona de cambium, unas pocas filas individuales de las cuales persisten como capas de formación celular durante algún tiempo. Durante los periodos de crecimiento, las células maduran continuamente a ambos lados del cambium, por lo que resulta bastante obvio que sólo una única capa de células puede tener existencia permanente como cambium.
Las otras capas, si están presentes, funcionan sólo temporalmente y se transforman completamente en células permanentes. En sentido estricto, sólo las iniciales constituyen el cambium, pero con frecuencia el término se utiliza con referencia a la zona cambial, porque es difícil distinguir las iniciales de sus derivados recientes.

Estructura celular del cambium:
Hay dos tipos diferentes de células de cambium:
1. Las iniciales del rayo, que son más o menos isodiamétricas y dan lugar a los rayos vasculares; y
2. Las iniciales fusiformes, las células cónicas alargadas que se dividen para formar todas las células del sistema vertical.
ADVERTENCIAS:
Las células cambiales están muy vacuoladas, normalmente con una gran vacuola y un citoplasma periférico delgado. El núcleo es grande y en las células fusiformes es muy alargado. Las paredes de las células cambiales tienen campos de fosas primarias con plasmodesmos. Las paredes radiales son más gruesas que las tangenciales y sus campos de fosas primarias están profundamente deprimidos.

División celular en el cambium:
Con el resultado de las divisiones tangenciales (periclinales) de las células del cambium se forman el floema y el xilema. Los tejidos vasculares se forman en dos direcciones opuestas, las células del xilema hacia el interior del eje, las del floema hacia su periferia. Las divisiones tangenciales de las iniciales del cambium durante la formación de los tejidos vasculares determinan la disposición de los derivados cambiales en filas radiales.
ADVERTENCIAS:
Como la división es tangencial, las células hijas que persisten como iniciales del cambium aumentan sólo en diámetro radial. Las nuevas iniciales del cambium formadas por divisiones transversales aumentan mucho en longitud; las formadas por divisiones radiales no aumentan en longitud.
A medida que el cilindro del xilema aumenta su grosor por el crecimiento secundario, el cilindro cambial también aumenta su circunferencia. La causa principal de este crecimiento es el aumento del número de células en dirección tangencial, seguido de una expansión tangencial de estas células.
Crecimiento del cámbium sobre las heridas:
Una de las funciones importantes del cambium es la formación de callo o tejido de las heridas, y la curación de las mismas. Cuando se producen heridas en las plantas, se forma una gran cantidad de tejido parenquimatoso blando sobre o debajo de la superficie lesionada; este tejido se conoce como callo. El callo se desarrolla a partir del cambium y por la división de las células del parénquima en el floema y la corteza.
Durante el proceso de curación de una herida se forma el callo. En éste hay al principio una abundante proliferación de las células del cambium, con la producción de parénquima masivo. Las células exteriores de este tejido se suberizan, o se desarrolla en ellas la peridermis, con lo que se forma una corteza.
Sin embargo, justo debajo de esta corteza el cambium permanece activo y forma nuevo tejido vascular de forma normal. El nuevo tejido formado de forma normal extiende la capa de crecimiento sobre la herida hasta que los dos lados opuestos se encuentran. Entonces las capas del cambium se unen y la herida queda completamente cubierta.
ADVERTENCIAS:
Cambium en brotes e injertos:
En las prácticas de brotación e injerto, el cambium tanto de la cepa como de la púa da lugar a un callo que se une y desarrolla una capa continua de cambium que da lugar al tejido conductor normal. Hay una unión real del cambium de la cepa y la púa de dos plantas durante las prácticas de brotación e injerto y, por lo tanto, estas prácticas no se encuentran comúnmente en las monocotiledóneas.
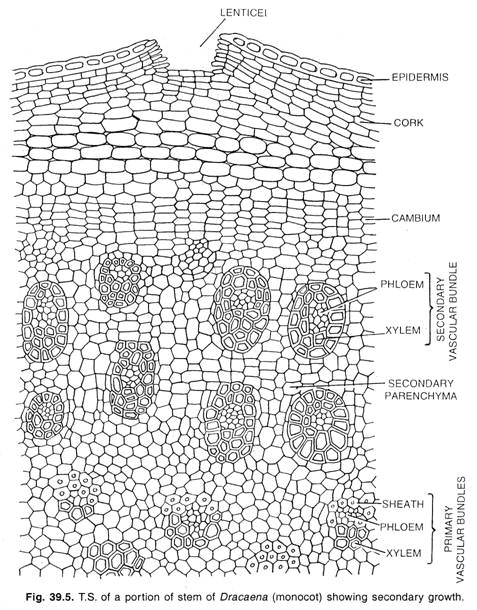
Cambium en Monocotiledóneas:
Un tipo especial de crecimiento secundario se da en pocas formas monocotiledóneas, como Dracaena, Aloe, Yucca, Veratrum y algunos otros géneros. En estas plantas el tallo aumenta de diámetro formando un cilindro de nuevos haces incrustados en un tejido.
Aquí se desarrolla una capa de cambium a partir del parénquima meristemático del peri-ciclo o de las células más internas de la corteza. En el caso de las raíces, el cambium de ésta se desarrolla en la endodermis. Las iniciales del cambium se encadenan en hileras para formar un cambium estratificado como el que se encuentra en el cambium normal de algunas dicotiledóneas.
Cambium en engrosamiento en las palmas:
ADVERTENCIAS:
Los tallos de las palmeras no aumentan su circunferencia, debido a ninguna actividad cambial, sino que este engrosamiento es el resultado del aumento gradual del tamaño de las células y de los espacios intercelulares y, a veces, de la proliferación de los tejidos fibrosos. Este es el tipo de crecimiento primario de larga duración.
El proceso es el siguiente:
La mayoría de las monocotiledóneas carecen de crecimiento secundario, pero con el resultado de un intenso y largo crecimiento primario continuado pueden producir cuerpos tan grandes como los de las palmeras. Las monocotiledóneas suelen producir un rápido engrosamiento por debajo del meristemo apical mediante un meristemo de engrosamiento primario periférico como se muestra en la figura.
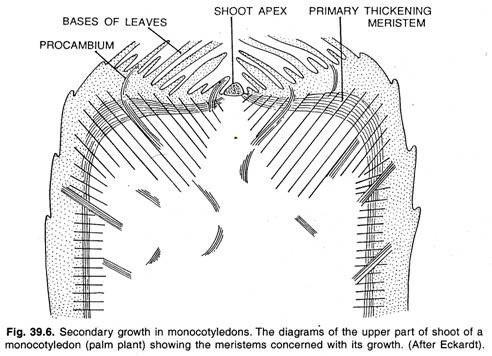
La actividad del meristemo de engrosamiento primario se asemeja con el crecimiento secundario encontrado en ciertas monocotiledóneas como Dracaena, Yucca, etc. El meristemo apical también conocido como ápice del brote produce sólo una pequeña parte del cuerpo primario, es decir, una columna central de parénquima y filamentos vasculares.
La mayor parte del cuerpo de la planta está formada por el meristemo primario de engrosamiento. El meristemo primario de engrosamiento se encuentra debajo del primordio de la hoja, que se divide periclinalmente produciendo hileras de células anticlinales. Estas células se diferencian en un tejido formado por un parénquima de fondo atravesado por filamentos procambiales.
Estos filamentos procambiales se convierten posteriormente en haces vasculares. Las células del parénquima basal se agrandan y dividen repetidamente, provocando un aumento de su grosor. De este modo, tanto el meristemo apical como el meristemo primario de engrosamiento dan lugar al grueso principal de los tejidos del tallo de las monocotiledóneas.
El engrosamiento se produce en las monocotiledóneas, como las palmeras, debido a las actividades del meristemo apical y del meristemo de engrosamiento primario.