ADVERTISEMENTS:
Impariamo il Cambio. Dopo aver letto questo articolo imparerai a conoscere: 1. Origine del cambio 2. Cambio fascicolare e interfascicolare 3. Durata 4. Funzioni 5. Struttura 6. Divisione cellulare 7. Ispessimento nelle palme.
Origine del cambio:
Lo scheletro vascolare primario è costruito dalla maturazione delle cellule dei filamenti di procambio per formare lo xilema e il floema. Nelle piante che non possiedono una crescita secondaria, tutte le cellule dei filamenti di procambio maturano e si sviluppano in tessuto vascolare.
Nelle piante che hanno una crescita secondaria in seguito, una parte dei filamenti di procambio rimane meristematica e dà origine al cambio vero e proprio. Nelle radici la formazione del cambium differisce da quella dei fusti a causa della disposizione radiale dei filamenti alternati di xilema e floema.
ADVERTISIMENTI:
Qui il cambio nasce come strisce discrete di tessuto nei filamenti di procambium all’interno dei gruppi di floema primario. Più tardi, le strisce di cambium con la loro estensione laterale si uniscono nel periciclo di fronte ai raggi dello xilema primario. La formazione del tessuto secondario è più rapida sotto i gruppi di floema così che il cambio, come si vede nella sezione trasversale delle radici più vecchie, forma presto un cerchio.
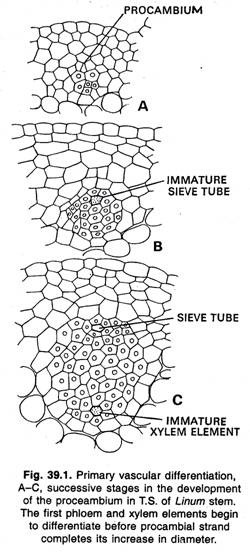
Cambium fascicolare e interfascicolare:
Nei fusti il primo procambio che si sviluppa dal promeristem si trova di solito sotto forma di filamenti isolati. In alcune piante questi primi filamenti formati diventano presto, uniti lateralmente da ulteriori filamenti simili formati tra loro e dall’estensione laterale dei primi filamenti formati.
Durante l’ulteriore sviluppo questo cilindro procambiale dà origine a un cilindro di tessuto vascolare primario (xilema e floema) e di cambium. Più tardi, si forma un cilindro di tessuto vascolare secondario che nasce in filamenti come il cilindro primario. In Ranunculus e in alcune altre piante erbacee, i filamenti di procambium, e i tessuti vascolari primari, non si fondono lateralmente ma rimangono come filamenti discreti.
ADVERTISIMENTI:
Più spesso nei fusti erbacei il cambium si estende lateralmente attraverso gli spazi intermedi fino a formare un cilindro completo. Dove si verifica tale estensione, il cambio nasce da cellule meristematiche interfascicolari derivate dal meristema apicale.
Le strisce di cambium che nascono all’interno dei fasci collaterali sono conosciute come cambium fascicolare, e le strisce cambiali che si trovano tra i fasci sono conosciute come cambium interfascicolare.
Durata del Cambio:
La durata della vita funzionale del cambium varia molto nelle diverse specie e anche nelle diverse parti della stessa pianta. In una pianta legnosa perenne il cambio del fusto principale vive dal momento della sua formazione fino alla morte della pianta.
È solo grazie alla continua attività del cambio nel produrre nuovo xilema e floema che tali piante possono mantenere la loro esistenza. Nelle foglie, nelle infiorescenze e in altre parti decidue, la vita funzionale del cambio è breve. Qui tutte le cellule del cambium maturano come tessuto vascolare. Lo xilema secondario si trova direttamente sul floema secondario in tali fasci.
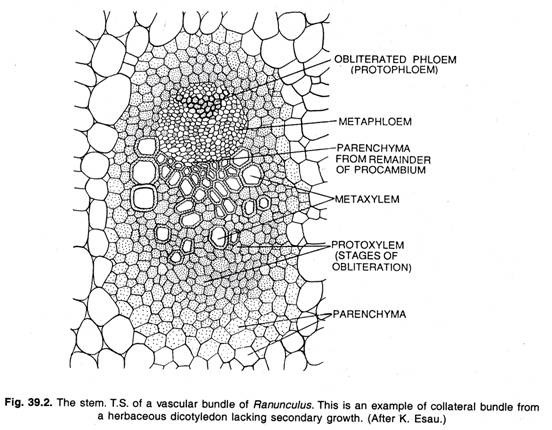
Funzione del Cambio:
Il meristema che forma i tessuti secondari consiste in un foglio uniseriato di iniziali che formano nuove cellule di solito su entrambi i lati. Il cambium forma lo xilema internamente e il floema esternamente. La divisione tangenziale della cellula cambiale forma due cellule figlie apparentemente identiche.
Una delle cellule figlie rimane meristematica, cioè la cellula cambiale persistente, l’altra diventa una cellula madre xilematica o una cellula madre floematica a seconda della sua posizione interna o esterna all’iniziale. La cellula cambiale si divide continuamente in modo simile; una cellula figlia rimane sempre meristematica, la cellula cambiale, mentre l’altra diventa o una cellula madre xilematica o una cellula madre floematica.
Probabilmente non c’è un’alternanza definita e per brevi periodi si forma un solo tipo di tessuto. Le cellule del cambium adiacenti si dividono quasi contemporaneamente e le cellule figlie appartengono allo stesso tessuto. In questo modo si mantiene la continuità tangenziale del cambio.
Struttura del Cambio:
ADVERTIME:
Ci sono due concezioni generali del cambium come strato iniziatore:
1. Che consiste in uno strato uniseriato di iniziatori permanenti con derivati che possono dividersi alcune volte e convertirsi presto in tessuto permanente;
2. Che ci sono diverse file di cellule iniziatrici che formano una zona di cambium, alcune singole file delle quali persistono come strati formanti cellule per qualche tempo. Durante i periodi di crescita le cellule maturano continuamente su entrambi i lati del cambio, diventa abbastanza ovvio che solo un singolo strato di cellule può avere esistenza permanente come cambio.
Gli altri strati, se presenti, funzionano solo temporaneamente e si trasformano completamente in cellule permanenti. In senso stretto, solo le iniziali costituiscono il cambium, ma spesso il termine è usato in riferimento alla zona cambiale, perché è difficile distinguere le iniziali dai loro derivati recenti.

Struttura cellulare del Cambio:
Ci sono due diversi tipi di cellule di cambium:
1. Le iniziali raggiate, che sono più o meno isodiametriche e danno origine ai raggi vascolari; e
2. Le iniziali fusiformi, le cellule affusolate allungate che si dividono per formare tutte le cellule del sistema verticale.
ADVERTIME:
Le cellule cambiali sono altamente vacuolate, di solito con un grande vacuolo e sottile citoplasma periferico. Il nucleo è grande e nelle cellule fusiformi è molto allungato. Le pareti delle cellule cambiali hanno campi di buche primarie con plasmodesmata. Le pareti radiali sono più spesse di quelle tangenziali, e i loro campi primari sono profondamente depressi.

Divisione cellulare nel cambio:
Con il risultato delle divisioni tangenziali (periclinali) delle cellule del cambium si formano il floema e lo xilema. I tessuti vascolari si formano in due direzioni opposte, le cellule dello xilema verso l’interno dell’asse, le cellule del floema verso la sua periferia. Le divisioni tangenziali delle iniziali del cambio durante la formazione dei tessuti vascolari determinano la disposizione dei derivati cambiali in file radiali.
ADVERTISIMENTI:
Siccome la divisione è tangenziale, le cellule figlie che persistono come iniziali del cambio aumentano solo di diametro radiale. Le nuove iniziali del cambio formate da divisioni trasversali aumentano molto in lunghezza; quelle formate da divisioni radiali non aumentano in lunghezza.
Come il cilindro xilematico aumenta di spessore per crescita secondaria, anche il cilindro cambiale cresce in circonferenza. La causa principale di questa crescita è l’aumento del numero di cellule in direzione tangenziale, seguito da un’espansione tangenziale di queste cellule.
Crescita del camice sulle ferite:
Una delle importanti funzioni del cambium è la formazione di callo o tessuto di ferita, e la guarigione delle ferite. Quando si verificano ferite sulle piante, una grande quantità di tessuto parenchimatoso morbido si forma su o sotto la superficie ferita; questo tessuto è noto come callo. Il callo si sviluppa dal cambium e dalla divisione delle cellule del parenchima nel floema e nella corteccia.
Durante il processo di guarigione di una ferita si forma il callo. In questo c’è all’inizio un’abbondante proliferazione delle cellule del cambium, con la produzione di un parenchima massiccio. Le cellule esterne di questo tessuto si suberizzano, o il periderma si sviluppa al loro interno, con il risultato che si forma una corteccia.
Tuttavia, proprio sotto questa corteccia il cambium rimane attivo e forma nuovo tessuto vascolare nel modo normale. Il nuovo tessuto formato nel modo normale estende lo strato di crescita sopra la ferita fino a quando i due lati opposti si incontrano. Gli strati di cambium si uniscono allora e la ferita viene completamente coperta.
ADVERTISIMENTI:
Cambium in Budding e Grafting:
Nelle pratiche di gemmazione e di innesto, il cambio sia del ceppo che della marza dà origine al callo che si unisce e sviluppa uno strato di cambio continuo che dà origine al tessuto conduttore normale. C’è un’effettiva unione del cambio del ceppo e della marza di due piante durante le pratiche di gemmazione e d’innesto e quindi queste pratiche non si trovano comunemente nelle monocotiledoni.
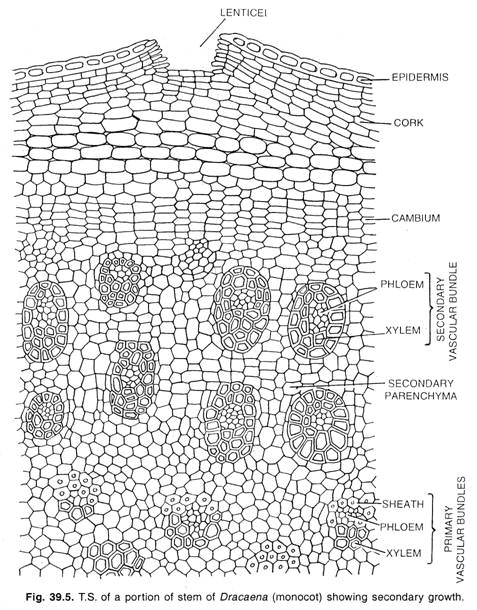
Cambium nelle monocotiledoni:
Un tipo speciale di crescita secondaria si verifica in poche forme monocotiledoni, come Dracaena, Aloe, Yucca, Veratrum e alcuni altri generi. In queste piante il fusto aumenta di diametro formando un cilindro di nuovi fasci incorporati in un tessuto.
Qui uno strato di cambium si sviluppa dal parenchima meristematico del peri-ciclo o dalle cellule più interne della corteccia. Nel caso delle radici, il cambium di questo si sviluppa nell’endoderma. Le iniziali del cambium si raggruppano in livelli per formare un cambium storiato come si trova nel cambium normale di alcune dicotiledoni.
Cambium in Thickening in Palms:
ADVERTISIMENTI:
I fusti delle palme non aumentano di circonferenza, a causa di una qualsiasi attività cambiale ma questo ispessimento è il risultato di un graduale aumento delle dimensioni delle cellule e degli spazi intercellulari e talvolta della proliferazione dei tessuti fibrosi. Questo è il tipo di crescita primaria di lunga durata.
Il processo è il seguente:
La maggior parte delle monocotiledoni manca di crescita secondaria, ma con il risultato di una crescita primaria intensa e prolungata possono produrre corpi così grandi come quelli delle palme. Le monocotiledoni spesso producono un rapido ispessimento sotto il meristema apicale per mezzo di un meristema primario periferico di ispessimento come mostrato in figura.
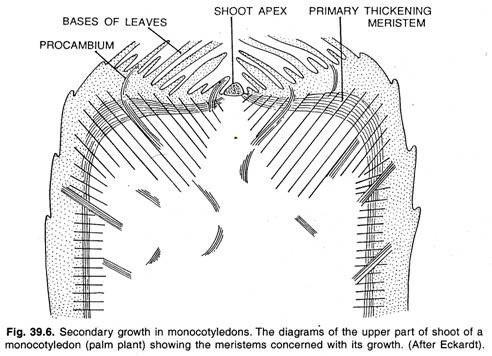
L’attività del meristema di ispessimento primario assomiglia alla crescita secondaria che si trova in certe monocotiledoni come Dracaena, Yucca, ecc. Il meristema apicale, noto anche come apice del germoglio, produce solo una piccola parte del corpo primario, cioè una colonna centrale di parenchima e filamenti vascolari.
La maggior parte del corpo della pianta è formata dal meristema primario di ispessimento. Il meristema primario di ispessimento si trova sotto la foglia-primordia, che si divide periclinalmente producendo file anticlinali di cellule. Queste cellule si differenziano in un tessuto formato da parenchima di terra attraversato da filamenti procambiali.
Questi filamenti procambiali si sviluppano in seguito in fasci vascolari. Le cellule del parenchima terrestre si ingrandiscono e si dividono ripetutamente, causando un aumento di spessore. In questo modo, sia il meristema apicale che il meristema primario di ispessimento danno origine alla maggior parte dei tessuti del fusto delle monocotiledoni.
L’ispessimento avviene nelle monocotiledoni, come le palme, a causa delle attività del meristema apicale e del meristema primario di ispessimento.