ADVERTISEMENTS:
Cambium について学びましょう。 この記事を読んだ後、あなたは以下のことを学びます。 1. 2. カンビウムの起源 筋膜と筋間膜 3. 持続時間 4. 機能 5. 構造 6. 細胞分裂 7. 手のひらの肥厚
カンビウムの起源。
一次的な血管骨格は、プロカンビウム筋の細胞が成熟して木部と葉茎を形成することにより構築される。 二次成長を持たない植物は、プロカンビウムストランドのすべての細胞が成熟し、血管組織に発展する。
後に二次成長を行う植物では、プロカンビウムストランドの一部が分裂性を保ち、本来のカンビウムを生じさせる。 根では木部と篩部が交互に放射状に配置されているため、根の形成は茎のそれとは異なる。
注意:
ここで子葉は、一次子葉のグループ内の子葉のストランドに組織の個別のストリップとして発生する。 その後、側方への伸長により、一次木部の光線に対向する周皮で接合される。 二次組織の形成は、葉茎群の下で最も速く、古い根の横断面を見ると、すぐに円形を形成する。
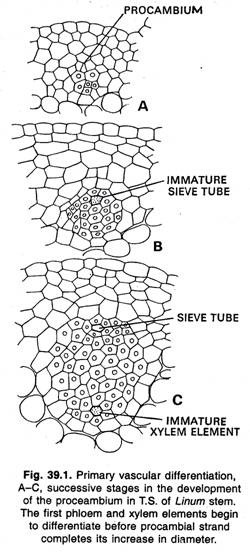
筋層および筋層間形成層。
茎では、前立茎から発生する最初の前腕は、通常、孤立したストランドの形で見られる。 いくつかの植物では、これらの最初に形成されたストランドは、それらの間に形成された追加の同様のストランドによって横方向に結合し、最初に形成されたストランドの横方向の伸長によってすぐになる。
さらに成長すると、この原始的な円柱は、第一次維管束組織(木部と篩部)と大脳皮質の円柱を生じます。 その後、二次維管束の円柱が形成され、一次維管束と同じように筋状に発生する。 ラナンキュラスや他の草本植物では、プロカンビウムのストランドと一次維管束組織は横方向に融合せず、個別のストランドとして残る。
注意事項:
草本の茎では、完全な円柱が形成されるまで、より多くの場合、子実体は間にある空間を横切って伸びる。 このような延長が起こる場合、幹細胞は頂端分裂組織から派生した筋間分裂細胞から発生する。
側枝束の中に生じた薄層は筋膜薄層と呼ばれ、束の間に見られる薄層は筋膜間薄層と呼ばれる。
上皮の長さ。
形成層が機能する期間は、種によって、また同じ植物でも部分によって大きく異なる。 多年生の木本植物では、主茎の形成時から植物の死までの間、子実体が生きている。
このような植物がその存在を維持できるのは、新しい木部と葉茎を作り出すという形成層の継続的な活動によってのみ可能なのである。 葉や花序などの落葉部では、大脳皮質の機能寿命は短い。 ここでは、すべての形成層が維管束組織として成熟する。 このような束では、二次木部は直接二次葉茎の上にある。
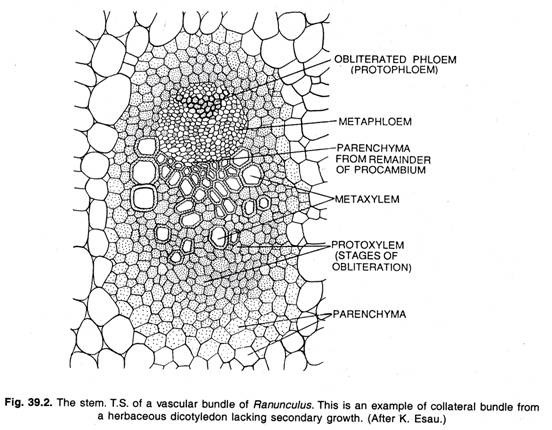
大脳皮質の機能:
二次組織を形成する分裂組織は、通常両側で新しい細胞を形成する初芽の単層シートで構成されています。 内部では木部、外部では葉茎を形成する。 子葉細胞の接線方向の分裂は、見かけ上同じ2つの娘細胞を形成する。
娘細胞の1つは分裂したままであり、すなわち根強い子葉細胞であり、もう1つは最初の内部または外部の位置に応じて木部母細胞または葉茎母細胞となる。 一方は木部母細胞、もう一方は葉茎母細胞になる。
おそらく明確な交替はなく、短い期間に一種類の組織だけが形成されるのであろう。 隣接するカンビウム細胞はほぼ同時に分裂し、娘細胞は同じ組織に属する。 このようにして、大脳皮質の接線方向の連続性が保たれる。
大脳皮質の構造。
注意事項:
開始層としての大脳皮質には2つの一般的な概念がある。
1. 2704>
1.永久的な初期細胞からなる単層で、数回分裂してすぐに永久的な組織となる誘導体がある。 2.初期細胞の列がいくつかあり、それが形成された層がしばらくの間持続するというもので、この層が形成された層が形成される。 成長期には、細胞は上皮の両側で連続的に成熟し、上皮として永続的に存在できるのは、単一の細胞層だけであることは明らかである。
他の層は、もし存在するならば、一時的にしか機能せず、完全に永久的な細胞へと変化していくのです。 厳密な意味では、初期のみが形成される。しかし、初期と最近の派生物を区別することは困難であるため、この用語はしばしば形成層と関連して使用される。

大脳皮質の細胞構造。
大脳皮質には2種類の細胞があります。
1.
1.光線原基:多かれ少なかれ等角的で、維管束線を生み出す。
2.楔状原基:細長い先細りの細胞で、分裂して垂直系の全細胞を形成する。
樟葉細胞は高度に空胞化しており、通常1つの大きな空胞と薄い周辺細胞質を持つ。 核は大きく、瘤状細胞ではかなり細長い。 壁には原始的なピットフィールドがあり、プラスモデスマータがある。 放射状の壁は接線方向の壁より厚く、一次ピットフィールドは深くくぼんでいる。

大脳皮質における細胞分裂:
接線方向の細胞分裂の結果、葉茎と木部が形成されます。 木部は軸の内側へ、葉部は軸の外側へと、それぞれ反対方向に形成される。 維管束形成時の子葉の接線方向の分裂が、子葉誘導体の放射状の列の配置を決定する。
ADVERTISEMENTS:
分割は接線方向なので、子細胞はcambium initialsとして存続し、径方向のみの直径を増加させる。 横方向の分裂で形成された新しい子実体は長さが大きく増加し、放射状の分裂で形成されたものは長さが増加しない。
木部円筒が二次成長によって厚みを増すと、子房円筒も円周が大きくなる。 この成長の主な原因は接線方向の細胞数の増加、次いでこれらの細胞の接線方向の膨張である。
傷口に関するカンビウムの成長。
カンビウムの重要な機能の1つは、カルスや傷の組織を形成し、傷を治すことです。 植物に傷ができると、傷の表面またはその下に軟らかい柔組織が大量に形成されるが、この組織をカルスという。 この組織はカルスと呼ばれる。カルスはカンビウムから、また篩骨と皮質の柔細胞が分裂することによって発生する。
傷が治る過程で、カルスが形成される。 このカルスでは、最初、多くの実質細胞が生成され、豊富な細胞増殖が見られる。 この組織の外側の細胞は硬くなり、その中に周皮が発生し、その結果樹皮が形成される。
しかし、この樹皮のすぐ下には大脳皮質が残っており、通常の方法で新しい血管組織が形成される。 通常の方法で形成された新しい組織は、傷の上に成長層を伸ばし、反対側の2つの側面が合わさるまで成長する。 その後、上皮の層が一体となり、傷は完全に覆われる。
注意事項:
芽かきと接ぎ木の際にカンビウムを使う。
芽かきと接ぎ木では、株と穂木の両方の子実体がカルスを生み、それが結合して連続した子実体層を形成し、正常な伝導組織を生み出します。 出芽や接ぎ木の際に、2つの植物の株と穂木の幹細胞が実際に結合するため、これらの方法は単子葉植物では一般的ではありません。
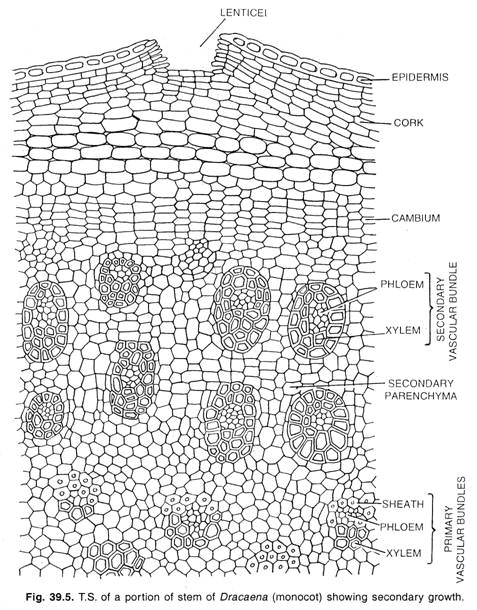
単子葉植物におけるカンビウム。
ドラセナ、アロエ、ユッカ、ベラチュラムなど、単子葉植物の一部には特殊な二次成長が見られます。 これらの植物では、茎は組織に埋め込まれた新しい束の円柱を形成して直径を増加させる。
ここで、周期の分裂柔細胞または皮質の最内部細胞から、形成されるのが、カンビウム層である。 根の場合、これの上皮は内皮に発達する。 一部の双子葉植物の正常な上皮に見られるように、上皮の初期が層状に連なり、層状の上皮を形成する。
ヤシの肥厚の中のカンビウム。
注意:
ヤシの茎が胴回りで増加するのは、いかなる形成層の活動によるものではなく、この肥厚は細胞のサイズと細胞間スペースが徐々に増加し、時には繊維組織の増殖の結果である。 これが長く続く一次成長のタイプである。
その過程は次のようなものである。
単子葉植物の多くは二次成長をしないが、一次成長が激しく長く続くと、ヤシの実のような大きな体を作ることができる。 単子葉植物は図に示すように、頂端分裂組織の下に周辺の一次肥厚分裂組織によって急速に肥厚することが多い。
一次肥厚分裂組織の活動は、ドラセナ、ユッカなどの単子葉植物に見られる二次成長に似ている。 シュートエーペックスとも呼ばれる頂端分裂組織は、原基のごく一部、すなわち柔細胞や維管束の中心柱しか作らない。
植物体の大部分は、一次肥厚分裂組織で形成される。 一次肥厚分裂組織は、葉原基の下にあり、周縁に分裂して反すうの細胞列を作る。 これらの細胞は、前子葉の筋が横切る地上柔組織を形成するように分化する。
これらの前子葉は、後に維管束に発展します。 地上実質細胞は肥大と分裂を繰り返し、厚みを増していく。 このようにして、頂端分裂組織と一次肥厚分裂組織の両方が、単子葉植物の茎組織の主要な大部分を生み出す。
ヤシなどの単子葉植物では、頂端分裂組織と一次肥厚分裂組織の活動によって肥大が起こる。