ADVERTISEMENTS:
Poznejme kambium. Po přečtení tohoto článku se dozvíte o: 1. Původ kambia 2. Fascikulární a interfascikulární kambium 3. Trvání 4. Funkce 5. Struktura 6. Dělení buněk 7. Tloušťkování v dlaních.
Původ kambia:
Primární cévní kostra se buduje zráním buněk prokambiových vláken za vzniku xylému a floému. U rostlin, které nemají sekundární růst, všechny buňky prokambiových pletiv dozrávají a vyvíjejí se v cévní tkáň.
U rostlin, které mají sekundární růst později, zůstává část prokambiových pletiv meristematická a vzniká z nich vlastní kambium. V kořenech se tvorba kambia liší od tvorby kambia ve stoncích kvůli radiálnímu uspořádání střídajících se vláken xylému a floému.
PŘEDPOKLADY:
Tady kambium vzniká jako diskrétní proužky tkáně v prokambiových vláknech uvnitř skupin primárního floému. Později se proužky kambia svým bočním rozšířením spojují v pericyklu naproti paprskům primárního xylému. Tvorba sekundárních tkání probíhá nejrychleji pod skupinami floému, takže kambium, jak je vidět na příčném řezu starších kořenů, brzy vytvoří kruh.
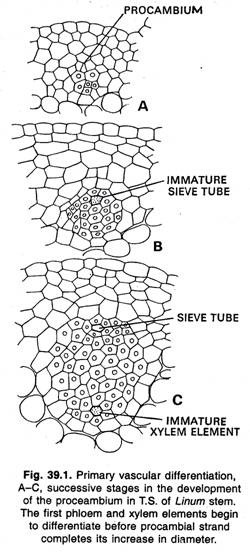
Fascikulární a interfascikulární kambium:
Ve stoncích se první prokambium, které se vyvíjí z promeristému, obvykle nachází ve formě izolovaných vláken. U některých rostlin se tato prvopramenná vlákna brzy spojují laterálně dalšími podobnými vlákny, která vznikají mezi nimi, a laterálním prodloužením prvopramenných vláken.
Během dalšího vývoje dává tento prokambiální válec vzniknout válci primárního cévního pletiva (xylému a floému) a kambii. Později vzniká válec sekundárního cévního pletiva, který vzniká v pramenech stejně jako primární válec. U pryskyřníku (Ranunculus) a některých dalších bylinných rostlin se prokambiové vlákno a primární cévní pletivo bočně nespojují, ale zůstávají jako oddělená vlákna.
DOPORUČENÍ:
Častěji se u bylinných stonků kambium rozšiřuje laterálně přes meziprostory, až se vytvoří úplný válec. Tam, kde k takovému rozšíření dochází, vzniká kambium z interfascikulárních meristematických buněk odvozených z apikálního meristému.
Pruhy kambia, které vznikají uvnitř kolaterálních svazků, se nazývají fascikulární kambium a pruhy kambia, které se nacházejí mezi svazky, se nazývají interfascikulární kambium.
Délka trvání kambia:
Délka funkčního života kambia se u různých druhů a také u různých částí téže rostliny značně liší. U vytrvalých dřevin žije kambium hlavního stonku od doby svého vzniku až do odumření rostliny.
Jedině díky pokračující činnosti kambia při tvorbě nového xylému a floému mohou takové rostliny udržovat svou existenci. V listech, květenstvích a dalších opadavých částech je funkční životnost kambia krátká. Zde všechny buňky kambia dozrávají jako cévní tkáň. Sekundární xylém se v takových svazcích nachází přímo na sekundárním floému.
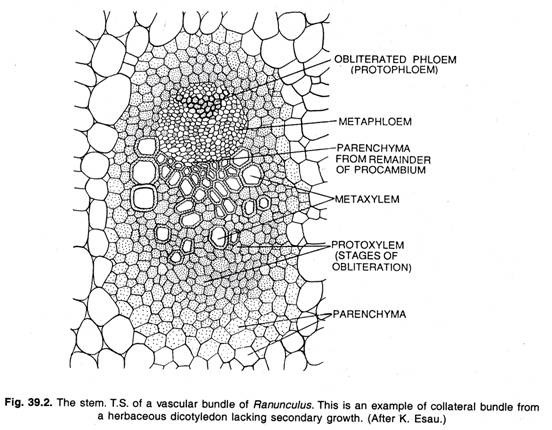
Funkce kambia:
Meristém, který tvoří sekundární pletiva, se skládá z jednovrstevného listu iniciál, které tvoří nové buňky obvykle na obou stranách. Kambium tvoří uvnitř xylém a navenek floém. Při tangenciálním dělení kambiální buňky vznikají dvě zdánlivě identické dceřiné buňky.
Jedna z dceřiných buněk zůstává meristematická, tj. trvalá kambiální buňka, druhá se stává xylémovou mateřskou buňkou nebo floémovou mateřskou buňkou v závislosti na své vnitřní nebo vnější poloze vůči iniciále. Podobným způsobem se průběžně dělí i buňka kambia; jedna dceřiná buňka vždy zůstává meristematická, tedy buňka kambia, zatímco druhá se stává buď xylémovou, nebo floémovou mateřskou buňkou.
Pravděpodobně nedochází k definitivnímu střídání a po krátká období se tvoří pouze jeden druh tkáně. Sousední buňky kambia se dělí téměř ve stejnou dobu a dceřiné buňky patří ke stejné tkáni. Tímto způsobem je zachována tangenciální kontinuita kambia.
Struktura kambia:
PŘEDPOKLADY:
Existují dvě obecné koncepce kambia jako iniciační vrstvy:
1. Že se skládá z jednovrstevné vrstvy trvalých iniciál s deriváty, které se mohou několikrát dělit a brzy se přeměnit v trvalé tkáně;
2. Že existuje několik řad iniciačních buněk, které tvoří kambiovou zónu, z nichž několik jednotlivých řad po určitou dobu přetrvává jako vrstvy tvořící buňky. Během vegetačního období dozrávají buňky průběžně na obou stranách kambia, je zcela zřejmé, že pouze jediná vrstva buněk může mít trvalou existenci jako kambium.
Další vrstvy, pokud jsou přítomny, fungují pouze dočasně a zcela se přemění na trvalé buňky. Ve striktním smyslu tvoří kambium pouze iniciály, ale často se tento termín používá s odkazem na kambiální zónu, protože je obtížné odlišit iniciály od jejich recentních derivátů.

Buněčná struktura kambia:
Existují dva různé typy buněk kambia:
1. Paprskovité iniciály, které jsou víceméně izodiametrické a dávají vznik cévním paprskům; a
2. Srostlé iniciály, podlouhlé zužující se buňky, které se dělí a vytvářejí všechny buňky vertikálního systému.
PŘÍČINY:
Kambiální buňky jsou silně vakuolizované, obvykle s jednou velkou vakuolou a řídkou periferní cytoplazmou. Jádro je velké a u srostlých buněk je značně protáhlé. Stěny kambiálních buněk mají primární jamková pole s plazmodesmaty. Radiální stěny jsou silnější než stěny tangenciální a jejich primární jamková pole jsou hluboce prohloubená.

Buněčné dělení v kambiích:
Výsledkem tangenciálního (periklinálního) dělení buněk kambia vznikají floém a xylém. Cévní pletiva se tvoří ve dvou opačných směrech, xylémové buňky směrem dovnitř osy, floémové buňky směrem k jejímu obvodu. Tangenciální dělení iniciál kambia při tvorbě cévních pletiv určuje uspořádání kambiálních derivátů v radiálních řadách.
DĚLENÍ:
Protože dělení je tangenciální, dceřiné buňky, které přetrvávají jako iniciály kambia, zvětšují pouze radiální průměr. Nové iniciály kambia vzniklé příčným dělením se výrazně zvětšují na délku; iniciály vzniklé radiálním dělením se na délku nezvětšují.
Jak xylémový válec sekundárním růstem zvětšuje svou tloušťku, zvětšuje se i obvod kambiálního válce. Hlavní příčinou tohoto růstu je zvýšení počtu buněk v tangenciálním směru a následné tangenciální rozšíření těchto buněk.
Růst kambia kolem ran:
Jednou z důležitých funkcí kambia je tvorba kalusu neboli ranného pletiva a hojení ran. Při poranění rostlin se na poraněném povrchu nebo pod ním vytvoří velké množství měkkého parenchymatózního pletiva; toto pletivo se nazývá kalus. Kalus se vyvíjí z kambia a dělením parenchymatických buněk ve floému a kůře.
Během procesu hojení rány se vytváří kalus. V něm dochází zpočátku k hojné proliferaci buněk kambia s tvorbou mohutného parenchymu. Vnější buňky tohoto pletiva se suberizují nebo se v nich vyvíjí periderm, v důsledku čehož vzniká kůra.
Těsně pod touto kůrou však kambium zůstává aktivní a normálním způsobem vytváří nové cévní pletivo. Nová tkáň vytvořená normálním způsobem rozšiřuje vegetační vrstvu přes ránu, dokud se obě protilehlé strany nesetkají. Vrstvy kambia se pak spojí a rána se zcela zakryje.
DOPORUČENÍ:
Kambium při pučení a roubování:
Při pupenování a roubování vzniká z kambia podnože i odnože kalus, který se spojuje a vytváří souvislou vrstvu kambia, z níž vzniká normální vodivé pletivo. Během postupů pupenování a roubování dochází ke skutečnému spojení kambia podnože a odnože dvou rostlin, a proto se tyto postupy běžně nevyskytují u jednoděložných rostlin.
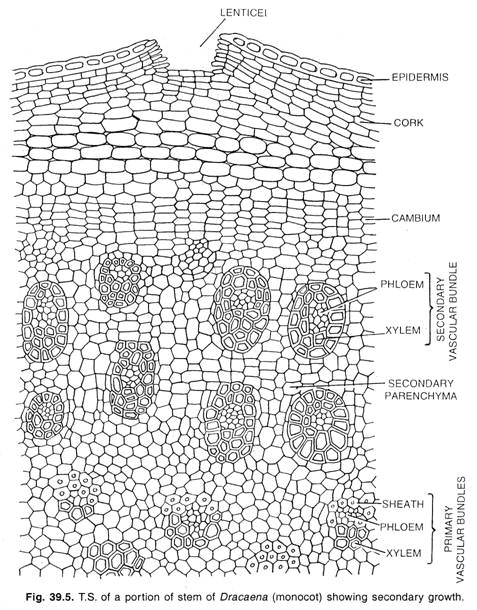
Kambium u jednoděložných rostlin:
Speciální typ sekundárního růstu se vyskytuje u několika jednoděložných forem, jako jsou Dracaena, Aloe, Yucca, Veratrum a některé další rody. U těchto rostlin se stonek zvětšuje v průměru a vytváří válec nových svazků uložených v pletivu.
Z meristematického parenchymu peri-cyklu nebo nejvnitřnějších buněk kůry se zde vyvíjí vrstva kambia. V případě kořenů se toto kambium vyvíjí v endodermis. Iniciály kambia se řetězí ve vrstvách a vytvářejí patrové kambium, jaké se vyskytuje v normálním kambium některých dvouděložných rostlin.
Kambium ve ztluštění u palem:
DOPORUČENÍ:
Stonky palem se nezvětšují v obvodu v důsledku jakékoli činnosti kambia, ale toto ztluštění je výsledkem postupného zvětšování velikosti buněk a mezibuněčných prostor a někdy i proliferace vláknitých pletiv. Jedná se o typ dlouho trvajícího primárního růstu.
Proces je následující:
Většina jednoděložných rostlin postrádá sekundární růst, ale v důsledku intenzivního a dlouho trvajícího primárního růstu mohou vytvářet tak velká těla, jako jsou těla palem. Jednoklíčníky často vytvářejí rychlé ztluštění pod apikálním meristémem pomocí periferního primárního ztlušťovacího meristému, jak je znázorněno na obrázku.
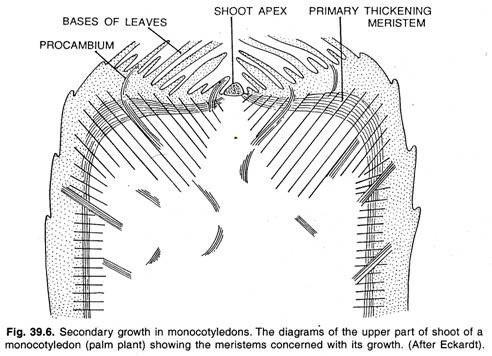
Činnost primárního ztlušťovacího meristému se podobá se sekundárním růstem, který se vyskytuje u některých jednoděložných rostlin, jako je Dracaena, Yucca atd. Apikální meristém známý také jako vrchol výhonu vytváří pouze malou část primárního těla, tj. centrální sloupec parenchymu a cévních vláken.
Většina rostlinného těla je tvořena primárním ztlušťujícím meristémem. Primární ztlušťující meristém se nachází pod listovým primordiem, které se dělí periklinálně a vytváří antiklinální řady buněk. Tyto buňky se diferencují v tkáň tvořenou přízemním parenchymem protkaným prokamenělými vlákny.
Tato prokambiální vlákna se později vyvíjejí v cévní svazky. Buňky přízemního parenchymu se zvětšují a opakovaně dělí, což způsobuje nárůst tloušťky. Tímto způsobem dává apikální meristém i primární ztlušťující meristém vzniknout hlavní části stonkových pletiv jednoděložných rostlin.
Ztlušťování probíhá u monokotyledonů, jako jsou palmy, díky činnosti apikálního meristému a primárního ztlušťovacího meristému.