ADVERTISERINGAR:
I den här artikeln kommer vi att diskutera svampar:- 1. Svamparnas historia 2. Definition av svampar 3. Ursprung 4. Allmänna kännetecken 5. Klassificering 6. Allmän organisation 7. Kanalsystem 8. Skelett 9. Reproduktion 10. Utveckling 11. Djurens natur 12. Affiniteter 13. Ekonomisk betydelse.
Innehåll:
- Svamparnas historia
- Definition av svampar
- Svamparnas ursprung
- Allmänt om svampar
- Klassificering av svampar
- Allmänt om svamparnas organisation
- Kanalsystem i
- Skelett hos svampar
- Rekonstruktion hos svampar
- Svampens utveckling
- Svampens djuriska karaktär
- Svampens egenskaper
- Svampens ekonomiska betydelse
Svampens historia:
Namnet Porifera (L., porus = por, ferro = bära) kommer från (1836). Svamparnas natur debatterades fram till långt in på 1800-talet, även om bevis för deras djuriska natur gavs 1765 av den som såg vattenströmmarna och rörelserna hos oscula. Som ett resultat av detta klassificerade Linné och Lamarck svamparna som zoofyter eller polyper och betraktade dem som besläktade med antozoiska coelenteraterater.
ADVERTISERINGAR:
Although (1816) separerade svamparna i en grupp Spongiaria allierad med Protozoa. Morfologin och fysiologin hos svamparna först förstods ordentligt av den som 1836 skapade namnet Porifera för den grupp som den nu är allmänt känd under. Iuxle (1875) och Sollas (1884) föreslog en fullständig separation av svamparna från andra Metazoa på grund av många särdrag.
Svamparna erkänns nu som en separat isolerad gren av Metazoa som efter Sollas kallas Parazoa.

Porifera innefattar svamparna som är de mest primitiva av de flercelliga djuren, de är fastsittande, växtliknande djur, de är fästa vid någon nedsänkt fast sten eller något fast skal och är oförmögna till någon rörelse. Porifera är uteslutande marina med undantag för en enda familj av sötvattensarter.
ÅTERKOMMELSER:
De kan ha cylindriska, grenade, vasliknande eller klotformiga former, vissa har en matt färg men de flesta har ljusa färger, de har röd, orange, lila, grön eller gul färg. Kroppen är perforerad av porer och kanaler men det finns inga organ, såsom mun eller nervsystem. Även om svampar är flercelliga djur bildar deras celler inte organiserade vävnader. De har vanligtvis ett endoskelett bestående av separata spikuler.
Förstörningen sker inom cellerna. På grund av sitt endoskelett och sina avskyvärda jäsningar äts de i allmänhet inte av djur. Svampar odlas för kommersiella ändamål.
Omkring 10 000 arter av svampar är för närvarande kända, och fylumet delas in i tre klasser, nämligen Calcarea eller Calcispongiae, Hexactinellida eller Hyalospongiae, och Demospongiae och omkring tolv ordningar huvudsakligen efter typen av skelett.
Definition av svampar:
Porifera kan definieras som ”asymmetriska eller radialsymmetriska flercelliga organismer med en cellulär organisationsgrad utan väldefinierade vävnader och organ; uteslutande vattenlevande; mestadels marina, sedentära, solitära eller koloniala djur med en kropp som är perforerad av porer, kanaler och kamrar genom vilka vatten strömmar; med ett eller flera inre hålrum som är fodrade med choanocyter; och med ett karakteristiskt skelett som består av kalksvampar, silikatiska spikuler eller horniga fibrer av spongin”.
Svampdjurens ursprung:
Det råder stor oenighet om Poriferas ursprung. De ser dock närmare ut som protozoer även om de skiljer sig från dem genom att vara flercelliga. Porifera har troligen sitt ursprung i flagellerade protozoer som Proterospongia, en kolonial flagellat. Kolonin av Proterospongia har celler med halsband och flageller som är inbäddade i en gelatinös matris med amöboida celler.
Dessa celler är svamparnas choanocyter och amöbocyter. Även då anses svamparnas ursprung vara osäkert. Man kan säga att svamparna skiljde sig från metazoernas evolutionära linje i ett mycket tidigt skede och att de därför har en separat status.
Allmänt om svampar:
1. Porifera är alla vattenlevande, mestadels marina utom en familj Spongillidae som lever i sötvatten.
2. De är fastsittande och fastsittande och växer som växter.
ADVERTIVERINGAR:
3. Kroppsformen är vas- eller cylinderliknande, asymmetrisk eller radiellt symmetrisk.
4. Kroppsytan är perforerad av många porer, ostia, genom vilka vattnet kommer in i kroppen och en eller flera stora öppningar, oscula, genom vilka vattnet passerar ut.
5. Flercellig kropp som består av yttre ektoderm och inre endoderm med ett mellanskikt av mesenkym, därför diploblastiska djur.

6. Kroppens inre utrymme är antingen ihåligt eller genomsyrat av många kanaler som kantas av choanocyter. Det inre utrymmet i svampkroppen kallas spongocoel.
ADVERTIVERINGAR:
7. Karakteristiskt skelett som består av antingen fina flexibla svampfibrer, kiselspikuler eller kalkspikuler.
8. Munnen saknas, matsmältningen är intracellulär, utsöndrings- och andningsorgan saknas.
9. Nerv- och känselcellerna är troligen inte differentierade.
ANNONSER:
10. Svamparna är enäggiga; fortplantning både genom asexuella och sexuella metoder.
11. Den asexuella förökningen sker genom knoppar och gemmules.
12. Svamparna besitter en hög regenerationskraft.
13. Den sexuella reproduktionen sker genom ägg och spermier; befruktningen är intern men korsbefruktning förekommer i regel.
ANNONSER:
14. Klyvning holoblastisk, utveckling indirekt genom en fritt simmande cilierad larv som kallas amfiblastula eller parenchymula.
15. Organisationen hos svampar har grupperats i tre huvudtyper, nämligen ascon-typ, sycon-typ och leuconoid-typ på grund av enkelhet i vissa former och komplexitet i andra.
Klassificering av svampar:
Klassificeringen av Porifera baseras huvudsakligen på de typer av skelett som finns hos dem. Detta fylum har klassificerats på olika sätt men den klassificering som föreslagits av Hyman (1940) och Burton (1967) är av stor betydelse. Den klassificering av Porifera som följs här är dock baserad på Storer och Usinger (1971) som verkar vara en modifierad form av Hymans klassificering.
Klass I. Calcarea (L., Calx = kalk) eller Calcispongiae:
(L., calx = kalk,Gr., spongos = svamp):
1. De har ett skelett av separata kalkspikuler som är monaxoniska eller tetraxoniska; tetraxoniska spikuler förlorar en stråle för att bli triadiska.
ADVERTISERINGAR:
2. De är solitära eller koloniala; kroppsformen är vasliknande eller cylindrisk.
3. De kan uppvisa asconoid, syconoid eller leuconoid struktur.
4. De är mattfärgade svampar som är mindre än 15 cm stora.
5. De förekommer i grunda vatten i alla hav.
Ordning 1. Homocoela:
1. Asconoida svampar med radialsymmetrisk cylindrisk kropp.
ADVERTIVERINGAR:
2. Kroppsväggen är tunn och inte veckad, spongocoel är fodrad av choanocyter.
Exempel:
Leucosolenia, Clathrina.
Ordning 2. Heterocoela:
1. Sykonoida eller leukonoida svampar med vasformad kropp.
2. Kroppsväggen är tjock och veckad, choanocyter kantar endast de radiella kanalerna.
3. Spongocoel är fodrad av tillplattade endodermceller.
Exempel:
Sycon eller Scypha, Grantia.
Klass. II. Hexactinellida eller Hyalospongiae:
(Gr., hyalos = glasig; spongos = svamp):
1. De kallas glassvampar; skelettet består av kiselhaltiga spikuler som är triaxonala med sex strålar. Hos vissa är spikulerna sammansmälta och bildar ett gitterliknande skelett.
2. Det finns inget epidermalt epitel.
3. Choanocyter kantar fingerformade kamrar.
4. De är cylindriska eller trattformade och finns i djupa tropiska hav, de blir upp till en meter.
Ordning 1. Hexasterophora:
1. Spikuler är hexastrar, det vill säga stjärnformade.
2. Radialkanaler eller flagellade kamrar är enkla.
3. De sitter inte fast med rotskotten utan är vanligen fästa vid ett hårt föremål.
Exempel:
Euplectella, Farnera.
Ordning 2. Amphidiscophora:
1. Spikuler är amfidiskar. Inga hexastrar.
2. De är knutna till substratet med rottofsar.
Exempel:
Hyalonema, Pheronema.
Klass III. Demospongiae (Gr., dermos = ram; spongos = svamp):
1. Den innehåller det största antalet svamparter. Storväxta, solitära eller koloniala.
2. Skelettet kan bestå av svampfibrer eller av svampfibrer med kiselalstrande spikuler eller så kan det saknas ett skelett.
3. Spikuler är aldrig sexstrålade, de är monaxon eller tetraxon och är differentierade i stora megaskleror och små mikroskleror.
4. Kroppens form är oregelbunden och kanalsystemet är av leucon-typ.
5. Generellt marina, få sötvattensformer.
Underklass I. Tetractinellida:
1. Svampar är mestadels fasta och enkla rundade kuddliknande tillplattade i formen vanligtvis utan grenar.
2. Skelettet består huvudsakligen av tetraxon-kiselspikuler men saknas i ordningen Myxospongida.
3. Kanalsystemet är av leukonoid typ. Form på grunt vatten.
Ordning 1. Myxospongida:
1. Enkel struktur.
2. Skelett saknas.
Exempel:
Oscarella, Halisarca.
Ordning 2. Carnosa:
1. Enkel struktur.
2. Spikuler är inte differentierade i megascleres och mikroscleres.
3. Aster kan förekomma.
Exempel:
Plakina.
Ordning 3. Choristida:
1. Spikuler differentieras i megascleres och mikroscleres.
Exempel:
Geodia, Thenea.
Subklass II. Monaxonida:
1. Monaxonider förekommer i olika former från rundad massa till förgrenad typ eller långsträckta eller stjälkformade med tratt eller fläktformade.
2. Skelettet består av monaxonidier med eller utan spongin.
3. Man skiljer på spikuler i megascleres och mikroscleres.
4. De finns i riklig mängd över hela världen.
5. Former på grunt och djupt vatten.
Ordning 1. Hadromerida:
1. Monaxon megascleres i form av tylostyler.
2. Mikrosklerer när de förekommer i form av aster.
3. Sponginfibrer saknas.
Exempel:
Cliona, Tethya.
Ordning 2. Halichondrida:
1. Monaxon megascleres är ofta av två typer, nämligen monactiner och diactiner.
2. Mikrosklerer saknas. Sponginfibrer förekommer men är sparsamma.
Exempel:
Halichondria.
Ordning 3. Poecilosclerida:
1. Monaxon megascleres är av två typer, en typ i ektodermen och en annan typ i choanocytskiktet.
2. Mikascleres är typiskt chelas, sigmas och toxas.
Exempel:
Cladorhiza.
Ordning 4. Haplosclerida:
1. Monaxon megascleres är av endast en typ, nämligen diactinal.
2. Mikrosklerer saknas.
3. Sponginfibrer är i allmänhet närvarande.
Exempel:
Chalina, Pachychalina, Spongilla
Underklass III. Keratosa:
1. Kroppen är rundad och massiv med ett antal iögonfallande oscula.
2. Skelettet består endast av ett nätverk av svampfibrer.
3. Det saknas kiselalger.
4. De är också kända som horniga svampar som finns i grunda och varma vatten i tropiska och subtropiska regioner.
Exempel:
Euspongia, Hippospongia.

Generell organisation av svampar:
Svampdjurens allmänna organisation varierar avsevärt. Svamparna är cylindriska som Leucosolenia, vasformade som Scypha och Grantia, trädliknande (t.ex. Microciona), fingerliknande (t.ex. Haliclona), bladliknande (t.ex. Phyllospongia), kuddeformade som Euspongia, repliknande (t.ex. Hyalonema), skålformade som Pheronema osv. Vissa svampar är solitära, medan andra är koloniala.
Svamparna är mestadels fastsittande former; de hittas fastsittande på stenar, snäckor, pinnar, sjögräs etc. Vissa är tråkiga svampar som Cliona. Vanligtvis är svampkroppen asymmetrisk, men några få former uppvisar radiell symmetri. Storleken varierar från några få mm till massiva med en diameter på 1 eller 2 meter.
Kroppsfärgningen hos svamparna varierar också mycket; de är oftast vita eller grå men gula, bruna, lila, orange, röda och gröna arter har också rapporterats. Svampens gröna färg beror vanligtvis på att det finns en symbiotisk alg, Zoo chlorella, i dem.

Kanalsystem hos svampar:
Alla håligheter i kroppen som genomkorsas av vattenströmmarna, som ger näring åt svampen från det att den tränger in genom porerna till dess att den passerar ut genom osculum, benämns gemensamt som kanalsystem. I Olynthus ses kanalsystemet i sin enklaste form.
I andra former kan det nå en hög grad av komplexitet, men dess allmänna utveckling kan ändå reduceras till en enkel tillväxtprocess hos den primitiva Olynthus som resulterar i veckning av väggarna och som åtföljs av en begränsning av de krageformade (choanocyt) cellerna till vissa regioner.
I den gradvisa och kontinuerliga differentieringsprocessen kan man urskilja tre distinkta typer av organisation som sammanlänkade genom många övergångar ändå kan betraktas som tre arkitekturstilar, så att säga under vilka alla existerande former kan klassificeras. Det finns vanligtvis tre typer av kanalsystem hos svampar, nämligen asconoid typ, syconoid typ och leuconoid typ.
(i) Asconoid typ:
Asconoid typ av kanalsystem är den enklaste av alla typer. I denna finns en radiellt symmetrisk vasliknande kropp som består av en tunn vägg som omsluter ett stort centralt hålrum spongocoel som öppnas i toppen av det förträngda osculum.
Väggen består av ett yttre och inre epitel med ett mesenchym emellan. Det yttre eller dermala epitelet som här kallas epidermis består av ett enda lager platta celler. Det inre epitelet, som kantar spongocoel, består av choanocyter. Mesenkymet innehåller skelettspikuler och flera typer av amöbocyter, alla inbäddade i en gelatinös matris.
Väggen i den asconoida svampen är perforerad av många mikroskopiska öppningar som kallas inkuranta porer eller ostior och som sträcker sig från den yttre ytan till spongocoeln. Varje por är intracellulär, dvs. det är en kanal genom en rörformig cell som kallas porocyt.

Vattenströmmen som drivs av choanocyternas flageller passerar genom de inkuranta porerna in i spongocoelen och ut genom osculum (vatten från utsidan → inkuranta porer → spongocoel → osculum → vatten ut) och förser den i sin passage med föda och syre och för bort metaboliska avfallsprodukter. Ett kanalsystem av asconoid typ finns endast hos ett fåtal svampar, t.ex. hos Olynthus och Leucosolenia.
Enligt Hyman är de viktiga kännetecknen för den asconoida strukturen den enkla väggen och det fullständiga kontinuerliga foderepitelet av choanocyter, som endast avbryts av de inre ändarna av porocyterna. Den asconoida typen av svamp liknar ytligt sett en typisk gastrula.
2. Sykonoid typ:
Sykonoid typ av kanalsystem är det första steget ovanför den askonoida typen. Det bildas genom att väggen i en asconoid svamp med jämna mellanrum trycks ut i fingerliknande utsprång, så kallade radiella kanaler.
I början är dessa radiella kanaler fria utsprång och det yttre vattnet omger hela deras längd, för det finns inga bestämda inkuranta kanaler. Men hos de flesta sykonoida svampar smälter radialkanalernas väggar samman på ett sådant sätt att de lämnar mellan sig rörformiga utrymmen, de inkuranta kanalerna, som öppnar sig utåt mellan de blinda yttre ändarna av radialkanalerna genom öppningar som kallas dermala ostior eller dermala porer.
Då dessa inkuranta kanaler utgör den ursprungliga yttre ytan hos den asconoida svampen är de med nödvändighet klädda med epidermis. Radialkanaler som är utskotten av det ursprungliga svampocoellet är nödvändigtvis fodrade av choanocyter och kallas därför bättre för flagellerade kanaler.
Det inre av den sykonoida svampen är ihåligt och bildar en stor spongocoel som är fodrad av det platta epitel som härrör från epidermis. De radiella kanalernas öppningar i spongocoelen kallas inre ostior. De sykonoida svamparna behåller askonoidernas radiella vasform och spongocoelen öppnar sig utåt genom ett enda terminalt osculum.
Väggen mellan de inkuranta och radiella kanalerna, är genomborrad av många små porer som kallas prosopyler. Vattenströmmen i sykonoida svampar tar följande väg dermala porer → incurrentkanaler → prosopyles → radiala kanaler → inre ostior (apopyles) → spongocoeL → osculum → ut.

De sykonoida svamparna skiljer sig från den askonoida typen på två viktiga punkter:
(i) I de tjocka vikta väggarna som innehåller omväxlande inkommande och radiella kanaler och
(ii) I brytningen av choanocytskiktet, som inte längre bekläder hela insidan utan är begränsat till vissa bestämda kamrar (radiella kanaler).

Den sykonoida strukturen uppträder i två huvudstadier. Den första typen illustreras i några få av de heterocoeliska kalksvamparna, särskilt medlemmar av släktet Sycon. I det andra stadiet sprider sig epidermis och mesenchym över den yttre ytan och bildar en tunn eller tjock cortex som ofta innehåller speciella kortikala spikuler. Epidermis blir genomborrad av mer bestämda porer som mynnar ut i förträngda incurrentkanaler.
3. Leukonoid typ:
Som ett resultat av den fortsatta processen med utvikning av choanocytskiktet och förtjockning av kroppsväggen utvecklas ett kanalsystem av leukonoid typ. Choanocytskiktet i den radiella kanalen i det sykonoida stadiet evagineras till många små kamrar, och dessa kan upprepa processen, så att kluster av små rundade eller ovala flagellerade kamrar ersätter de långsträckta kamrarna i det sykonoida stadiet.
Koanocyterna är begränsade till dessa kamrar. Mesenchym fyller ut utrymmena runt de flagellformade kamrarna. Spongocoelen är vanligtvis utplånad och hela svampen blir oregelbunden i sin struktur och obestämd i sin form. Svampens inre genomsyras av många inkommande och utgående kanaler som förenas till större utgående kanaler och utrymmen som leder till oscula.
Överytan är täckt av epidermalt epitel och är genomborrad av många dermala porer (ostia) och oscula.
De dermala porerna leder till incurrentkanaler som förgrenar sig oregelbundet genom mesenkymet. De inkuranta kanalerna leder in i de små rundade flagellade kamrarna genom öppningar som fortfarande benämns prosopyle. De flagellformade kamrarna öppnar sig genom öppningar som kallas apopyler till ex-current kanaler, och dessa förenas till större och större rör, av vilka de största leder till oscula.

De viktigaste kännetecknen för den leukonoida typen av kanalsystem är choanocyternas begränsning till små kamrar, den stora utvecklingen av mesenkymet och komplexiteten hos de inkommande och utkommande kanalerna.
Vattenströmmens förlopp är dermal ostia → incurrent canals → prosodus (om det finns) → prosopyles → flagellated chambers → apopyles → aphodus (om det finns) → excurrent canals → större kanaler → oscula → out. Kanalsystemet av leucon-typ är mycket effektivt och de flesta svampar är byggda enligt den leuconoida planen och de uppnår en avsevärd storlek.
De är alltid oregelbundna till sin struktur men flödet av vattenströmmen är ganska snabbt och effektivt. Den leuconoida typen av kanalsystem uppvisar många variationer men presenterar tre utvecklingsstadier, nämligen eurypylous, aphodal och diplodal.
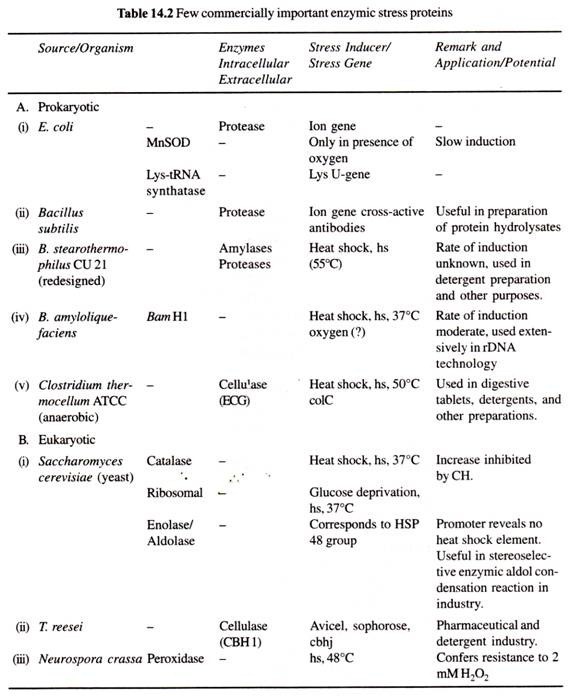
(a) Eurypylous:
I det eurypyloida leuconoida kanalsystemet är de flagellerade kamrarna breda och fingerborgsformade, var och en öppnar sig direkt i den exkursiva kanalen genom en bred öppning som kallas apopyle och tar emot vattentillförseln direkt från den inkursiva kanalen genom prosopyle.
Vattenströmmen tar följande väg → dermala porer eller ostior → subdermala utrymmen → incurrent canals → prosopyles → flagellated chambers → apopyles -> excurrent canals spongocoel → oscula → out. Denna typ av kanalsystem finns hos Leucilla.
(b) Aphodal:
I den aphodala leuconoida typen av kanalsystem är de flagellerade kamrarna små och rundade. Öppningen från varje flagellad kammare till den exkursiva kanalen dras ut i ett smalt rör, vanligtvis inte särskilt långt, som kallas aphodus. Förhållandet mellan de flagellerade kamrarna och de inkuranta kanalerna förblir som tidigare.
Vattenströmmens väg är följande dermala porer eller ostior → subdermalt utrymme → incurrent canals → prosopyles → flagellated kamrar → aphodus → excurrent canals→ spongocoel → oscula → out. Denna typ av kanalsystem finns hos Geodia och Stelleta.
(c) Diplodal:
I vissa fall finns det också ett smalt strömningsrör, prosodus, mellan den inkuranta kanalen och flagellkamrarna, ett sådant tillstånd kallas diplodal. Denna typ av kanalsystem finns bland annat hos Oscarella och Spongilla.
Vattenströmmen tar följande väg:
Dermala porer eller ostior → subdermala utrymmen → inkuranta kanaler → prosodus → flagellated kamrar → aphodus → exkuranta kanaler → spongocoel → oscula → ut.
4. Rhagon typ:
I kalkhaltiga svampar kan den leukonoida strukturen uppnås genom askonoida och sykonoida stadier. Men hos Demospongiae härstammar den från ett stadium som kallas rhagon, som i sin tur uppstår genom direkt omläggning av den inre cellmassan.

Rhagontypen av svamp har en bred bas och den är konisk till formen med ett enda osculum i toppen. Den basala väggen benämns hypophare som saknar flagellerade kamrar. Den övre väggen som bär en rad små, ovala flagellerade kamrar kallas spongophare.
Spongocoel är avgränsad av ovala flagellerade kamrar som mynnar ut i den genom breda apopyler. Mellan kamrarna och epidermis ligger en betydande tjocklek av mesenkym som genomkorsas av incurrentkanaler och subdermala utrymmen. Dermala porer eller ostior mynnar ut i subdermala utrymmen som sträcker sig under hela kroppens yta.
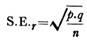
Breddande incurrentkanaler leder från de subdermala utrymmena in i små flagellerade kamrar som har bildats genom nedbrytning av radialkanalen, de flagellerade kamrarna är ensamma fodrade av choanocyter. Från de flagellerade kamrarna leder ex-current canals in i en spongocoel.
De incurrenta och ex-currenta kanalerna kan vara komplexa och förgrenade. Spongocoelen öppnas genom ett enda osculum. Vattenströmmens lopp är ostia→ subdermala utrymmen → incurrent canals → prosopyles → flagellated chambers → apopyles → ex-current canals → spongocoel → osculum → out.
Kanalsystemets funktioner:
Kanalsystemet hjälper svamparna med näring, andning, utsöndring och reproduktion. Vattenströmmen som flyter genom kanalsystemet för med sig näring och syre och för bort koldioxid, kvävehaltigt avfall och avföring.
Den transporterar spermier från en svamp till en annan för befruktning av äggcellerna. Kanalsystemet ökar också svamparnas yta i kontakt med vattnet och gör det därmed möjligt för svamparna att öka sin volym eftersom förhållandet mellan yta och volym måste förbli konstant.
Skelett i svampar:
Enligt Haeckel har få svampar som bor i djuphavet ett pseudoskelett som helt består av främmande kroppar utan några element som utsöndras av svampen själv. Den stora majoriteten av svamparna har dock ett verkligt skelett som kallas autoskelett och som består av element som utsöndras av svampen själv. Auto skelett i svampar är antingen spikuler eller spongin eller en kombination av båda.

Reproduktion hos svampar:
Svamparna förökar sig både asexuellt och sexuellt.
1. Asexuell reproduktion:
Akönad fortplantning förekommer i alla Porifera. Den sker genom regenerering, bildning av reduktionskroppar, knoppbildning och gemmulebildning.
(i) Regeneration:
Kraften till regeneration är mycket stor hos svampar, vilken del som helst som skärs av kan regenerera hela svampen. Om svampen macereras och pressas genom en fin silkesduk kommer dess celler och cellkluster att passera igenom, dessa kan regenerera nya svampar. Regenereringsförmågan används industriellt för odling av badsvampar.
(ii) Bildning av reduktionskroppar:
En annan mycket ovanlig metod för asexuell reproduktion är bildandet av reduktionskroppar.
Många sötvattens- och havssvampar faller sönder under ogynnsamma omständigheter. Den sönderfallande svampen kollapsar vanligtvis och lämnar små rundade bollar, så kallade reduktionskroppar. Varje reduktionskropp består av en inre massa amöbocyter, som utvändigt täcks av en pinacoderm. När de gynnsamma förhållandena återkommer växer dessa reduktionskroppar till helt nya svampar.
(iii) Knoppbildning:
I svampar sker knoppbildning på olika sätt.
(a) Exogen knoppbildning:
En svamp bildar yttre knoppar vegetativt vid basen av grenar och bildar på så sätt en grupp individer. Så småningom drar knopparna ihop sig från föräldern och var och en bildar en ny svamp.
(b) Endogen knoppbildning:
Akutana fortplantningskroppar som kallas gemmules bildas internt hos alla sötvattenssvampar och vissa havssvampar.
Arkeocyter samlas i grupper i mesogloea, de blir då flerkärniga, de fylls också med proteiner som reservnäringsmaterial som tillförs arkeocyterna av speciella ammaceller trophocyter. Vissa amöbocyter omsluter denna massa av matfyllda arkeocyter och utsöndrar ett hårt dubbelskiktat skal, skalet har ett litet utlopp eller mikropyle.
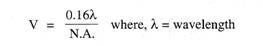
Därefter utsöndrar vissa skleroblaster spikuler som är placerade radiellt mellan två lager av skalet, vissa spikuler sticker ut utanför skalet, spikulerna i Spongilla är monaxonspikuler men hos andra är de amfidisker. Amfidiskerna är raka stavar med taggiga sidor och en ring av krokar i varje ände.
Så bildas en gemmula varefter de omgivande amöbocyterna, skleroblasterna och trophocyterna avgår. Gemmules bildas i stort antal på hösten varefter svampen sönderfaller, de stannar kvar i svampresterna eller blir fria, i vilket fall som helst faller de till botten. Gemmules kan klara ogynnsamma förhållanden, men de kläcks när våren kommer.
Vid kläckningen strömmar arkeocyterna ut ur mikropylen, sedan delar sig dessa flerkärniga arkeocyter och bildar icke-kärniga arkeocyter och små celler som kallas histoblaster.
Histoblasterna bildar genom differentiering och omorganisering epidermis porocyter, choanocyter och inre endodermfoder; modifierade arkeocyter från skleroblast som utsöndrar spikler. Ungefär en vecka efter kläckningen omger en ung svamp det tomma gemmulosskalet. De okärniga arkeocyterna förblir embryonala.

2. Sexuell reproduktion:
Svampar har inga könsorgan men amöbocyter bildar könsceller i mesenkymet ; först produceras ägg och senare spermier, därför är svampen protogyn och korsbefruktning äger rum. Amöbocyterna fylls med mat och blir stora, de blir runda och bildar ägg.
Andra amöboocyter delar sig för att producera ett stort antal spermier, en spermie har ett ovalt huvud och en lång avsmalnande svans.
Vissa arbetare hävdar att könsceller bildas från arkeocyter eller till och med från choanocyter. Ägg från en svamp befruktas av spermier från en annan svamp för att bilda zygoter. Vid korsbefruktning går spermien troligen in i en choanocyt som smälter samman med ägget och frigör spermien som sedan smälter samman med ägget. Zygoten utsöndrar ett hölje som kallas yngelkapsel och som omsluter zygoten.

Svampdjurens utveckling:
Zygoten genomgår en holoblastisk men ojämlik klyvning; vid holoblastisk delning är zygoten helt segmenterad.
De tre första delningarna är vertikala som bildar åtta pyramidala celler, därefter bildar en horisontell delning åtta små övre celler vid djurpolen och åtta stora nedre celler vid växtpolen. De övre små cellerna delar sig snabbt, blir tydliga och får flageller, medan de nedre cellerna delar sig långsamt och blir korniga.
Detta bildar en blastula som har ett blastocoelhålrum invändigt. Blastula kallas amfiblastula efter bildandet av flageller. Hittills har utvecklingen skett i svampens kropp, nu letar sig amfiblastula in i ex-currentkanaler och lämnar föräldern genom osculum.
Amfiblastula simmar omkring fritt i några timmar, sedan invagineras de övre flagellerade cellerna i blastocoel och de växer över av de nedre granulära cellerna; detta bildar en gastrula som är som en kopp med ett yttre skikt av granulära celler som kallas ektoderm, och ett inre skikt av flagellerade celler som kallas endoderm, den har en stor öppning, blastoporen.
Detta embryo fäster sig vid något fast föremål med sin blastoporala ände och det börjar växa. Både ektoderm- och endodermskikten utsöndrar mesogloea och dess amöbocyter. Enligt vissa utsöndras mesogloea endast av choanocyter (endoderm). Väggen förtjockas och viks till kanaler, perforeringar bildar ostia och osculum.

Kimskikten hos svampar är inte likvärdiga med ektoderm och endoderm hos Metazoa, eftersom svampens yttre ektoderm har bildats av de nedre granulära cellerna i växtpolen, och endodermen bildas av de övre flagellerade cellerna i djurpolen. I Metazoa blir cellerna i djurpolen ektoderm och cellerna i växtpolen bildar endoderm.
Svamparnas animaliska natur:
Svampar betraktades länge inte som djur, just på grund av att de inte ser ut som djur; de är fastsittande och har ingen förmåga att fånga föda eller göra sig av med sitt avfall och reagerar mer eller mindre inte på stimuli.
Det var dock Robert Grant (1836) som erkände svamparnas djuriska karaktär genom att föreslå att vattenströmmen flyter genom en bestämd väg i svampkroppen; den kommer in i svampkroppen genom små porer, så kallade ostior, som är utspridda över hela kroppsytan, och passerar ut genom större hål, så kallade oscula.
Svamparnas holozoiska näringssätt; de livnär sig på fasta matpartiklar som följer med vattenströmmen, saknar cellulosacellvägg, och närvaron av frisimmande ciliatlarver i livscykeln som liknar andra havsdjurs livscykler tyder tydligt på att svamparna är djur och inget annat.
Affiniteter hos svampar:
Det har funnits en stor kontrovers om svampdjurens natur och släktskap ända sedan de upptäcktes. Aristoteles (384-322 f.Kr.)var den förste som erkände dem som djur. Men biologerna efter Aristoteles trodde i århundraden att de var växter på grund av deras fastsittande form och okänsliga natur.
Ellis (1765) påpekade att en kontinuerlig vattenström går in i och ut ur svampkroppen, vilket bekräftades av Robert Grant (1836). Linné, Lamarck och Cuvier placerade dem dock tillsammans med coelenteraterater i Zoophyta. Robert Grant (1836) fastställde slutligen en separat grupp Porifera för svamparna. De är alltså besläktade med både protozoer och metazoer.
Affiniteter med protozoer:
Likheter:
Likheter:
Svampar uppvisar likheter med koloniala protozoer genom att de har följande egenskaper:
1. Avsaknad av matsmältningshålor och förekomst av intracellulär matsmältning.
2. Förekomst av krageceller och amöboida celler som hos koloniala flagellater.
3. Produktion av skelett av en enskild cell eller en grupp av celler.
4. Cellerna i svampkroppen är beroende av varandra i sin funktion.
5. En process som kallas inversion sker i amfiblastula-larver som de av Volvox. De flagellerade svamplarverna anses vara rester av någon flagellerad kolonial protozoisk förfader.
Unlikheter:
Svampar skiljer sig från protozoer genom att de har följande egenskaper:
1. Kanalsystemet.
2. Det karakteristiska skelettet.
3. Utvecklingen av en flercellig kropp från ett befruktat ägg genom klyvning, och genom denna egenskap blir svamparna mer komplexa än koloniala protozoer och liknar metazoerna. Därför är dess protozoiska släktskap inte längre giltigt.
Affiniteter med Metazoa:
Liknande:
Av Metazoa uppvisar coelenteraterna vissa likheter med svamparna i följande egenskaper:
1. Båda är sedentära till sin vana.
2. Båda är diploblastiska och acoelomater. Ett icke-cellulärt mesogloea-skikt finns mellan ektoderm och endoderm. Mesoderm saknas i båda fallen.
3. Svampdjurens spongocoel som öppnar sig genom det terminala osculum kan jämföras med det gastrokärlska hålrummet som öppnar sig utåt genom det terminala hypos tome hos coelenteraterater.
4. Asexuell reproduktion sker i båda fallen och kolonierna bildas genom knoppbildning.
5. Parenchymula-larven hos svampar är jämförbar med planula hos coelenteraterater.
Underskotten:
Svamparna skiljer sig från Metazoa genom att de har följande egenskaper:
1. Hos svamparna är cellerna jämförelsevis mindre specialiserade och beroende av varandra än cellerna hos Metazoa.
2. Svampar har choanocyter som inte finns hos någon Metazoa.
3. Svampar har en cellulär uppbyggnad, medan Metazoa har en vävnads- och organuppbyggnad.
4. Svamparna är diploblastiska, medan Metazoa (utom Coelenterata) är triploblastiska.
5. Svamparna har inte en främre ände eller ett huvud som Metazoa har. Osculum fungerar endast som utandningsöppning.
5. Svamparnas kropp är perforerad av många små porer och de har ett unikt system av kanaler i sin kropp; alla dessa finns aldrig hos Metazoa.
6. Svampar har inga nervvävnader för att samordna kroppens aktiviteter.
7. Det saknas helt och hållet matsmältningshålor hos svamparna, till skillnad från dem hos Metazoa.
8. Svampar har ett större antal fetter och fettsyror som har en högre molekylvikt än de som finns hos Metazoa.
9. Vid utvecklingen av svampar blir embryots flagellerade övre celler, belägna vid djurpolen, till endoderm och de granulära cellerna vid växtpolen blir till ektoderm; hos ingen Metazoa sker en sådan inversion under utvecklingen.
10. På grund av dessa skillnader kan svamparna inte grupperas med Metazoa och därför har ett separat fylum skapats för svamparna.
Till trots sina likheter med Protozoa och Metazoa tror man alltså att svamparna har avvikit från huvudlinjen i utvecklingen av Metazoa och att de är ett fylum i en återvändsgränd. Med tanke på detta har ett separat underrike Parazoa skapats för att inkludera svamparna och därför har de en isolerad fylogenetisk position.
Svampdjurens ekonomiska betydelse:
Svamparna har stor ekonomisk betydelse, både till nytta och till skada för mänskligheten.
A. Nyttiga svampar:
Svamparna är nyttiga för människan och andra djur på följande sätt:
1. Som föda:
Svampar tas sällan till föda av andra djur på grund av deras dåliga smak, lukt och vassa taggar. Kräftdjur lever dock som parasiter på dem och vissa blötdjur, t.ex. nakensnäckor, är beroende av dem som föda.
2. Som kommensaler:
Svampar fungerar som skyddshus för flera djur som kräftdjur, maskar, blötdjur, småfiskar etc., eftersom deras fiender inte kan äta svamparna. Förutom skyddet får de djur som lever inuti svampkroppen ett rikt födoämne från det vatten som cirkulerar genom dem. Dessa djur orsakar inte någon uppenbar skada på svamparna, de betraktas som kommensaler.
3. Andra användningsområden:
Det torkade, fibrösa skelettet av många svampar som Spongia, Hippospongia och Euspongia används för att bada, polera, tvätta bilar, väggar, möbler, skrubba golv osv. Skelettet av vissa svampar, t.ex. Euplectella, har ett stort kommersiellt värde och används som dekorativa föremål.
B. Skadliga svampar:
Bara ett fåtal svampar är skadliga. De kan orsaka döden för vissa fastsittande djur genom att växa över dem och skära av deras mat- och syretillgång. De tråkiga svamparna, som Cliona, fäster sig vid skalen på ostron, musslor, havstulpaner osv. Den borrar sig in i dessa djurs skal och förstör dem helt och hållet.
De tråkiga svamparna orsakar också stor skada på ostronbottnar. De borrande svamparna förstör också stenar genom att tränga in i dem och bryta dem i bitar.
