ADVERTISEMENTS:
I denne artikel vil vi diskutere svampe:- 1. Svampenes historie 2. Definition af svampe 3. Oprindelse 4. Generelle karakterer 5. Klassifikation 6. Generel organisation 7. Kanalsystem 8. Skelet 9. Reproduktion 10. Udvikling 11. Dyrenes natur 12. Affiniteter 13. Økonomisk betydning.
Indhold:
- Svampenes historie
- Definition af svampe
- Svampenes oprindelse
- Generelle karakterer af svampe
- Klassifikation af svampe
- Svampenes generelle organisation
- Kanalsystem i Svampe
- Skelet hos svampe
- Reproduktion hos svampe
- Svampes udvikling
- Svampes dyriske natur
- Affiniteter hos svampe
- Svampes økonomiske betydning
Svampes historie:
Navnet Porifera (L., porus = pore; ferro = at bære) stammer fra (1836). Svampenes natur blev diskuteret indtil langt ind i det nittende århundrede, selv om beviset for deres dyriske natur blev givet i 1765 af den, der så vandstrømmene og bevægelserne i oscula. Som følge heraf klassificerede Linné, Lamarck og klassificerede svampene under zoofytter eller polypper og betragtede dem som beslægtede med antozoiske coelenteraterater.
ADVERTISEMENTS:
Although (1816) adskilte svampene i en gruppe Spongiaria, der var beslægtet med protozoer. Svampenes morfologi og fysiologi blev først tilstrækkeligt forstået af, der i 1836 skabte navnet Porifera for den gruppe, som den nu er almindeligt kendt under, iuxle (1875) og Sollas (1884) foreslog en fuldstændig adskillelse af svampene fra de andre Metazoa på grund af mange særheder.
Svampe anerkendes nu som værende en særskilt isoleret gren af Metazoa, der efter Sollas kaldes Parazoa.

Porifera omfatter svampene, som er de mest primitive af de flercellede dyr, de er fastsiddende, planteagtige dyr, de er fastgjort til en eller anden nedsænket fast sten eller skal og er ude af stand til at bevæge sig. Porifera er udelukkende marine med undtagelse af en enkelt familie af ferskvandsarter.
ADVERTISEMENTS:
Deres form kan være cylindrisk, forgrenet,vaseformet eller kugleformet, nogle er matte i farven, men de fleste er farvestrålende, de har rød, orange, lilla, grøn eller gul farve. Kroppen er perforeret af porer og kanaler, men der er ingen organer, som f.eks. mund eller nervesystem. Selv om svampe er flercellede dyr, danner deres celler ikke organiserede væv. De har normalt et endoskelet bestående af separate spicula.
Fuldstændig fordøjelse finder sted inden i cellerne. På grund af deres endoskelet og ubehagelige gæringer bliver de generelt ikke spist af dyr. Svampe dyrkes til kommercielle formål.
Omkring 10.000 arter af svampe er kendt på nuværende tidspunkt, og stamtræet er opdelt i tre klasser, nemlig Calcarea eller Calcispongiae, Hexactinellida eller Hyalospongiae og Demospongiae og omkring tolv ordener hovedsageligt efter skelet-typen.
Definition af svampe:
Porifera kan defineres som “asymmetriske eller radialt symmetriske flercellede organismer med en cellulær organisationsgrad uden veldefinerede væv og organer; udelukkende akvatiske; for det meste marine, sedentære, solitære eller koloniale dyr med en krop perforeret af porer, kanaler og kamre, som vandet strømmer igennem; med et eller flere indre hulrum foret med choanocytter; og med et karakteristisk skelet af kalkholdige spicula, kiselholdige spicula eller hornede sponginfibre”.
Svampernes oprindelse:
Der er stor uenighed om oprindelsen af Porifera. De ligner dog nærmere protozoer, selv om de adskiller sig fra dem ved at være flercellede. Porifera stammer sandsynligvis fra flagellede protozoer som Proterospongia, der er en koloni-flagellat. Proterospongia-kolonien består af celler med krave og flageller, der er indlejret i en geléagtig matrix med amøboide celler.
Disse celler er svampenes choanocytter og amoebocytter. Selv da menes det, at svampenes oprindelse er usikker. Det kan konkluderes, at svampene adskilte sig fra metazoernes udviklingslinje på et meget tidligt tidspunkt og derfor har en særskilt status.
Generelle karakterer af svampe:
1. Porifera er alle akvatiske, for det meste marine med undtagelse af en familie Spongillidae, som lever i ferskvand.
2. De er siddende og fastsiddende og vokser som planter.
ADVERTISERINGER:
3. Kropsformen er vase- eller cylinderformet, asymmetrisk eller radialt symmetrisk.
4. Kroppens overflade er perforeret af talrige porer, ostia, hvorigennem vandet kommer ind i kroppen, og en eller flere store åbninger, oscula, hvorigennem vandet passerer ud.
5. Flercellet legeme bestående af ydre ektoderm og indre endoderm med et mellemliggende lag af mesenchym, derfor diploblastiske dyr.

6. Kroppens indre rum er enten hult eller gennemtrængt af talrige kanaler, der er foret med choanocytter. Det indre rum i svampekroppen kaldes spongocoel.
ADVERTISER:
7. Karakteristisk skelet bestående af enten fine fleksible svampefibre, kiselholdige spicula eller kalkholdige spicula.
8. Mund mangler, fordøjelsen er intracellulær; ekskretions- og åndedrætsorganer mangler.
9. Nerve- og sansecellerne er sandsynligvis ikke differentierede.
ANNONCER:
10. Svampene er enboede; formering både ved aseksuelle og seksuelle metoder.
11. Den aseksuelle formering sker ved knopper og gemmoler.
12. Svampene besidder en høj regenerationsevne.
13. Den kønslige formering sker ved hjælp af æg og sædceller; befrugtningen er intern, men krydsbefrugtning forekommer som regel.
ANNONCER:
14. Spaltning holoblastisk, udvikling indirekte gennem en fritsvømmende cilieret larve kaldet amfiblastula eller parenchymula.
15. Organiseringen af svampe er blevet inddelt i tre hovedtyper, nemlig ascon-typen, sycon-typen og leuconoid-typen på grund af enkelhed i nogle former og kompleksitet i andre.
Klassifikation af svampe:
Klassifikationen af Porifera er hovedsageligt baseret på de typer af skeletter, der findes hos dem. Dette phylum er blevet klassificeret forskelligt, men den klassifikation, som Hyman (1940) og Burton (1967) har foreslået, er af betydelig betydning. Den klassifikation af Porifera, der følges her, er imidlertid baseret på Storer og Usinger (1971), som synes at være en modificeret form af Hyman’s klassifikation.
Klasse I. Calcarea (L., Calx = kalk) eller Calcispongiae:
(L., calx = kalk,Gr., spongos = svamp):
1. De har et skelet af adskilte kalkspicula, der er monaxon eller tetraxon; tetraxon-spicula mister en stråle og bliver triadiate.
ADVERTISERINGER:
2. De er solitære eller koloniale; kropsformen er vaselignende eller cylinderformet.
3. De kan udvise asconoid, syconoid eller leuconoid struktur.
4. De er matte farvede svampe, der er mindre end 15 cm store.
5. De forekommer i lavt vand i alle oceaner.
Ordning 1. Homocoela:
1. Asconoide svampe med radialsymmetrisk cylinderformet krop.
ADVERTISER:
2. Kropsvæggen er tynd og ikke foldet, svampocoel er beklædt af choanocytter.
Eksempler:
Leucosolenia, Clathrina.
Ordning 2. Heterocoela:
1. Sykonoide eller leukonoide svampe med vaseformet krop.
2. Kropsvæggen er tyk og foldet, choanocytterne beklæder kun radialkanalerne.
3. Svampeskallen er beklædt af fladtrykte endodermceller.
Eksempler:
Sycon eller Scypha, Grantia.
Klasse. II. Hexactinellida eller Hyalospongiae:
(Gr., hyalos = glasagtig; spongos = svamp):
1. De kaldes glassvampe; skelettet er af kiselholdige spicules, som er triaxoniske med seks stråler. Hos nogle er spiculerne smeltet sammen og danner et gitteragtigt skelet.
2. Der er ikke noget epidermisk epithel.
3. Choanocytter beklæder fingerformede kamre.
4. De er cylindriske eller tragtformede og findes i dybe tropiske have, de bliver op til en meter.
Ordning 1. Hexasterophora:
1. Spicules er hexastere, dvs. stjernelignende i form.
2. Radiale kanaler eller flagellerede kamre er simple.
3. De er ikke fæstnet med rodskud, men er almindeligvis fæstnet til en hård genstand.
Eksempler:
Euplectella, Farnera.
Ordning 2. Amphidiscophora:
1. Spicules er amphidiscus. Ingen hexasters.
2. De er fastgjort til substratet med rottetufter.
Eksempler:
Hyalonema, Pheronema.
Klasse III. Demospongiae (Gr., dermos = ramme; spongos = svamp):
1. Den indeholder det største antal svampearter. Store størrelser, solitære eller koloniale.
2. Skelettet kan bestå af svampefibre eller af svampefibre med kiselholdige spikuler, eller der kan ikke være noget skelet.
3. Spicula er aldrig sekstråede, de er monaxoniske eller tetraxoniske og er differentieret i store megasclerer og små mikrosclerer.
4. Kropsformen er uregelmæssig, og kanalsystemet er af leucon-typen.
5. Generelt marine, få ferskvandsformer.
Underklasse I. Tetractinellida:
1. Svampe er for det meste faste og enkle afrundede afrundede pudeformede fladtrykte i form normalt uden forgreninger.
2. Skelettet består hovedsageligt af tetraxon-kiselholdige spicula, men mangler i ordenen Myxospongida.
3. Kanalsystemet er af leukonoid type. Lavvandsform.
Ordning 1. Myxospongida:
1. Enkel struktur.
2. Skelet mangler.
Eksempler:
Oscarella, Halisarca.
Ordning 2. Carnosa:
1. Enkel struktur.
2. Spicules er ikke differentieret i megascleres og mikroscleres.
3. Der kan være aster til stede.
Eksempel:
Plakina.
Ordning 3. Choristida:
1. Spicules er differentieret i megascleres og mikroscleres.
Eksempler:
Geodia, Thenea.
Underklasse II. Monaxonida:
1. Monaxonider forekommer i forskellige former fra afrundet masse til forgrenet type eller langstrakt eller stilkede med tragt eller vifteformet.
2. Skelettet består af monaxonidier med eller uden spongin.
3. Spiculer skelnes i megascleres og mikroscleres.
4. De findes i rigelige mængder i hele verden.
5. Lavvandede og dybe vandformer.
Ordning 1. Hadromerida:
1. Monaxon megascleres i form af tylostyler.
2. Mikrosklerer, når de findes i form af aster.
3. Sponginfibre er fraværende.
Eksempler:
Cliona, Tethya.
Ordning 2. Halichondrida:
1. Monaxon megascleres er ofte af to typer, nemlig monactiner og diactiner.
2. Mikrosklerer er fraværende. Sponginfibre er til stede, men sparsomt fordelt.
Eksempel:
Halichondria.
Ordning 3. Poecilosclerida:
1. Monaxon megascleres er af to typer, en type i ektodermen og en anden type i choanocytlaget.
2. Mikroscleres er typisk chelas, sigmas og toxas.
Eksempel:
Cladorhiza.
Ordning 4. Haplosclerida:
1. Monaxon megascleres er kun af én type, nemlig diactinal.
2. Mikrosklerer er fraværende.
3. Sponginfibre er generelt til stede.
Eksempler:
Chalina, Pachychalina, Spongilla
Underklasse III. Keratosa:
1. Kroppen er afrundet og massiv med en række iøjnefaldende oscula.
2. Skelet består kun af et net af svampefibre.
3. Kiselholdige spicula er fraværende.
4. De er også kendt som hornede svampe, der findes i lavt og varmt vand i tropiske og subtropiske områder.
Eksempler:
Euspongia, Hippospongia.

Generel organisering af svampe:
Den generelle organisation af svampe varierer betydeligt. Svampene er cylindriske som Leucosolenia, vaseformede som Scypha og Grantia, trælignende (f.eks. Microciona), fingerlignende (f.eks. Haliclona), fingerlignende (f.eks. Haliclona), bladlignende (f.eks. Phyllospongia), pudeformede som Euspongia, tovformede (f.eks. Hyalonema), skålformede som Pheronema osv. Nogle svampe er solitære, mens andre er koloniale.
Svampene er for det meste fastsiddende former; de findes fastgjort til sten, skaller, pinde, havgræs osv.; nogle er kedelige svampe som Cliona. Normalt er svampekroppen asymmetrisk, men få former udviser radialsymmetri, men få former udviser radialsymmetri. Størrelsen varierer fra få mm til massive svampe med en diameter på 1 eller 2 meter.
Svampernes kropsfarve varierer også meget; de er for det meste hvide eller grå, men der er også rapporteret gule, brune, lilla, orange, røde og grønfarvede arter. Svampens grønne farve skyldes normalt tilstedeværelsen af en symbiotisk alge, Zoo chlorella, i dem.

Kanalsystem hos svampe:
Alle de hulrum i kroppen, der gennemstrømmes af de vandstrømme, som nærer svampen, fra den kommer ind gennem porerne, til den passerer ud gennem osculum, betegnes samlet som kanalsystem. Hos Olynthus ses kanalsystemet i sin enkleste form.
I andre former kan det nå en høj grad af kompleksitet, men dets generelle udvikling kan ikke desto mindre reduceres til en simpel vækstproces hos den primitive Olynthus, der resulterer i en foldning af væggene og ledsages af en begrænsning af de kraveformede (choanocyt) celler til visse områder.
I den gradvise og kontinuerlige differentieringsproces kan der skelnes mellem tre forskellige organisationstyper, som forbundet af talrige overgange alligevel kan betragtes som tre byggestilarter, under hvilke alle eksisterende former så at sige kan klassificeres. Der findes normalt tre typer af kanalsystemer hos svampe, nemlig den asconoide type, den syconoide type og den leuconoide type.
(i) Asconoid type:
Asconoid type af kanalsystem er den enkleste af alle typer. I denne er der et radialt symmetrisk vaseformet legeme bestående af en tynd væg, der omslutter et stort centralt hulrum svampeskallen, der åbner sig i toppen ved det indsnævrede osculum.
Væggen er sammensat af et ydre og indre epitel med et mesenchym imellem. Det ydre eller dermale epithel, her kaldet epidermis, består af et enkelt lag af flade celler. Det indre epithel, der beklæder spongocoelet, består af choanocytter. Mesenchymet indeholder skeletspicula og flere typer amøbocytter, der alle er indlejret i en geléagtig matrix.
Væggen i den asconoide svamp er perforeret af talrige mikroskopiske åbninger, der betegnes incursive porer eller ostier, som strækker sig fra den ydre overflade til spongocoelen. Hver pore er intracellulær, dvs. det er en kanal gennem en rørformet celle kaldet en porocyt.

Vandstrømmen, der drives af choanocytternes flageller, passerer gennem de indgående porer ind i svampeskallen og ud gennem osculum (vand fra ydersiden → indgående porer → svampeskallen → osculum → vand ud) og forsyner på sin vej føde og ilt og fører metaboliske affaldsstoffer bort. Asconoid type af kanalsystem findes kun hos få svampe, f.eks. hos Olynthus og Leucosolenia.
Ifølge Hyman er de vigtige kendetegn ved den asconoide struktur den simple væg og det fuldstændige sammenhængende foringsepitel af choanocytter, der kun afbrydes af de indre ender af porocytterne. Den asconoide svampetype ligner overfladisk set en typisk gastrula.
2. Sykonoid type:
Sykonoid type af kanalsystem er det første stadium over den askonoide type. Det er dannet ved, at væggen af en asconoid svamp med regelmæssige mellemrum skubbes ud i fingerlignende fremspring, kaldet radiale kanaler.
I begyndelsen er disse radiale kanaler frie fremspring, og det ydre vand omgiver hele deres længde, for der er ingen deciderede indgående kanaler. Men hos de fleste sykonoide svampe smelter radialkanalernes vægge sammen paa en saadan maade, at der imellem dem er rørformede rum, de indstrømmende kanaler, som åbner sig udadtil mellem de blinde ydre ender af radialkanalerne ved aabninger, der kaldes dermal ostia eller dermalporer.
Da disse indgående kanaler udgør den oprindelige ydre overflade af den asconoide svamp, er de nødvendigvis beklædt med epidermis. Radiale kanaler, der er udposninger af den oprindelige svampeskal, er nødvendigvis beklædt med choanocytter og kaldes derfor bedre flagelkanaler.
Det indre af den sykonoide svamp er hult og danner et stort svampeskrog, som er foret med det flade epitel, der stammer fra epidermis. De radiale kanalers åbninger ind i svampeskallen betegnes som indre ostier. De sykonoide svampe har bevaret asconoidernes radiale vaseform, og svampeskallen åbner sig udadtil ved det enkelte terminale osculum.
Væggen mellem de indgående og de radiale kanaler, er gennemboret af talrige små porer kaldet prosopyler. Vandstrømmen i sykonoide svampe tager følgende rute dermale porer → incurrentkanaler → prosopyles → radiale kanaler → indre ostier (apopyles) → spongocoeL → osculum → ud.

De sykonoide svampe adskiller sig fra den askonoide type på to vigtige punkter:
(i) ved de tykke foldede vægge, der indeholder skiftevis indgående og radiale kanaler, og
(ii) ved afbrydelsen af choanocytlaget, der ikke længere beklæder hele det indre, men er begrænset til visse bestemte kamre (radiale kanaler).

Den sykonoide struktur optræder i to hovedstadier. Den første type illustreres hos nogle få af de heterocoeløse kalksvampe, især hos medlemmer af slægten Sycon. I det andet stadium spreder epidermis og mesenchym sig over den ydre overflade og danner en tynd eller tyk cortex, der ofte indeholder særlige kortikale spicula. Epidermis bliver gennemboret af mere tydelige porer, som munder ud i indsnævrede indgående kanaler.
3. Leuconoid type:
Som følge af en yderligere proces med udfoldning af choanocytlaget og fortykkelse af kropsvæggen udvikles den leukonoide type af kanalsystem. Choanocytlaget i radialkanalen i det syconoide stadie udsvinder i mange små kamre, og disse kan gentage processen, således at klynger af små afrundede eller ovale flagellede kamre erstatter de langstrakte kamre i det syconoide stadie.
Koanocytterne er begrænset til disse kamre. Mesenchym fylder rummene omkring de flagellede kamre ud. Svampeskallen er normalt udslettet, og hele svampen bliver uregelmæssig i sin struktur og ubestemmelig i sin form. Svampens indre bliver gennemsyret af mange indgående og udgående kanaler, der går sammen og danner større udgående kanaler og rum, som fører til oscula.
Overfladen er dækket af epidermal epithelium og er gennemboret af mange dermale porer (ostia) og oscula.
De dermale porer munder ud i incurrentkanaler, der forgrener sig uregelmæssigt gennem mesenchymet. De indgående kanaler munder ud i de små afrundede flagellede kamre ved åbninger, der stadig betegnes prosopyler. De flagellede kamre åbner sig ved åbninger, der kaldes apopyler, til ex-current-kanaler, og disse forenes til større og større rør, hvoraf de største fører til oscula.

De vigtigste kendetegn ved den leukonoide type af kanalsystem er choanocytternes begrænsning til små kamre, den store udvikling af mesenchymet og kompleksiteten af de indstrømmende og udstrømmende kanaler.
Vandstrømmens forløb er dermal ostia → incurrente kanaler → prosodus (hvis til stede) → prosopyles → prosopyles → flagelkamre → apopyles → aphodus (hvis til stede) → excurrente kanaler → større kanaler → oscula → ud. Kanalsystemet af leucon-typen er meget effektivt, og de fleste svampe er bygget efter den leuconoide plan, og de opnår en betydelig størrelse.
De er altid uregelmæssigt opbygget, men strømmen af vandets strøm er ret hurtig og effektiv. Den leuconoide type af kanalsystem udviser talrige variationer, men præsenterer tre udviklingsstadier, nemlig eurypylous, aphodal og diplodal.
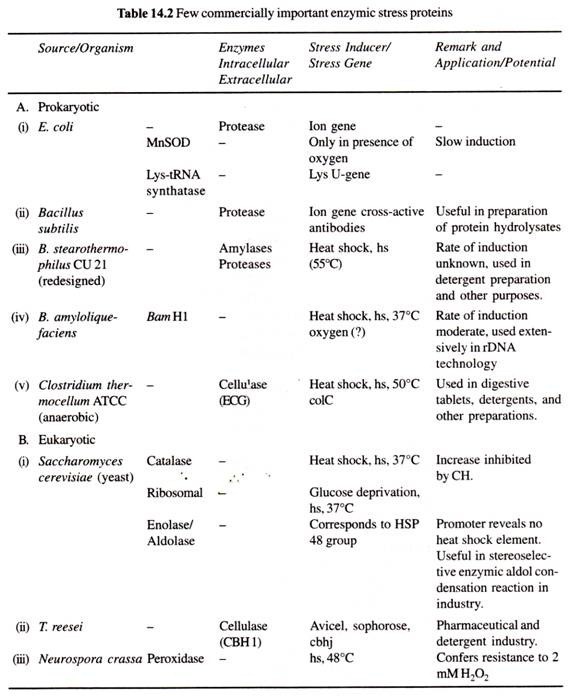
(a) Eurypylous:
I det eurypyloide leuconoide kanalsystem er de flagellede kamre brede og fingerbølformede, idet de hver især åbner direkte ind i den udstrømmende kanal ved en bred åbning kaldet apopyle og modtager vandtilførslen direkte fra den indstrømmende kanal gennem prosopyle.
Vandstrømmen tager følgende rute → dermale porer eller ostier → subdermale rum → incurrentkanaler → prosopyle → flagelkamre → apopyle -> excurrentkanaler spongocoel → oscula → ud. Denne type kanalsystem findes hos Leucilla.
(b) Aphodal:
I den aphodale leuconoide type af kanalsystem er de flagellerede kamre små og afrundede. Åbningen fra hvert flagelkammer ind i den ekskurrerende kanal er trukket ud i et smalt rør, som regel ikke er særlig langt, kaldet aphodus. Flagelkamrenes forhold til de indgående kanaler forbliver som før.
Vandstrømmens rute er som følger dermale porer eller ostier → subdermalt rum → incurrentkanaler → prosopyles → flagelkamre → aphodus → excurrentkanaler→ spongocoel → oscula → ud. Denne type kanalsystem findes hos Geodia og Stelleta.
(c) Diplodal:
I nogle tilfælde er der også et smalt strømrør, prosodus, mellem den incurbide kanal og de flagellede kamre, en sådan tilstand kaldes diplodal. Denne type af kanalsystem findes hos Oscarella, Spongilla osv.
Vandstrømmen tager følgende rute:
Dermale porer eller ostier → subdermale rum → indadgående kanaler → prosodus → flagelerede kamre → aphodus → udadgående kanaler → svampocoel → oscula → udad.
4. Rhagon type:
I kalkholdige svampe kan den leuconoide struktur opnås ved hjælp af asconoide og syconoide stadier. Men hos Demospongiae er den afledt af et stadium kaldet rhagon, som igen opstår ved direkte omlægning af den indre cellemasse.

Rhagon-typen af svamp har en bred base, og den er kegleformet med et enkelt osculum på toppen. Den basale væg betegnes hypophare, som er blottet for flagellerede kamre. Den øverste væg med en række små, ovale flagellede kamre kaldes spongophare.
Spongocoel er afgrænset af ovale flagellede kamre, der munder ud i den ved brede apopler. Mellem kamrene og epidermis ligger en betydelig tykkelse af mesenchym, der gennemskæres af indgående kanaler og subdermale rum. Dermale porer eller åbninger munder ud i subdermale rum, der strækker sig under hele kroppens overflade.
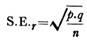
Gennemgående incurrerende kanaler fører fra de subdermale rum ind i små flagellede kamre, der er blevet dannet ved nedbrydning af radialkanalen, de flagellede kamre er alene foret med choanocytter. Fra de flagellede kamre fører udstrømmende kanaler ind i en svampeskal.
De incurrente og ex-currente kanaler kan være komplekse og forgrenede. Spongocoelen åbner sig ved et enkelt osculum. Vandets strømningsforløb er ostia→ subdermale rum → incurrentkanaler → prosopyler → flagellede kamre → apopyler → ex-currentkanaler → spongocoel → osculum → ud.
Funktioner af kanalsystemet:
Kanalsystemet hjælper svampene med ernæring, respiration, udskillelse og reproduktion. Den vandstrøm, der strømmer gennem kanalsystemet, bringer føde og ilt med sig og fjerner kuldioxid, kvælstofholdigt affald og afføring.
Det transporterer sædcellerne fra den ene svamp til den anden med henblik på befrugtning af æggene. Kanalsystemet øger også svampenes overfladeareal i kontakt med vandet og gør det således muligt for svampene at øge deres volumen, da overfladevolumenforholdet skal forblive fast.
Skelet i svampe:
I henhold til Haeckel har få svampe, der lever i dybhavet, et pseudo skelet, der udelukkende består af fremmedlegemer uden nogen elementer udskilt af svampen selv. Langt de fleste svampe besidder derimod et ægte skelet kaldet auto skelet, der består af elementer udskilt af svampen selv. Auto-skelettet i svampe er enten spicules eller spongin eller en kombination af begge dele.

Fremstilling hos svampe:
Svampene formerer sig både aseksuelt og seksuelt.
1. Aseksuel reproduktion:
Aseksuel formering forekommer overalt hos Porifera. Den finder sted ved regeneration, dannelse af reduktionslegemer, knopdannelse og gemmedannelse.
(i) Regeneration:
Fornyelsesevnen er meget stor hos svampene, enhver afskåret del vil regenerere hele svampen. Hvis svampen udblødes og presses gennem et fint silketørklæde, vil dens celler og celleklumper passere igennem, disse kan regenerere nye svampe. Regenereringskraften anvendes industrielt til dyrkning af badesvampe.
(ii) Dannelse af reduktionslegemer:
En anden meget usædvanlig metode til aseksuel formering er dannelsen af reduktionslegemer.
Mange ferskvands- og havsvampe går i opløsning under ugunstige omstændigheder. Den opløste svamp vil normalt falde sammen og efterlade små afrundede kugler, kaldet reduktionslegemer. Hvert reduktionslegeme består af en indre masse af amoebocytter, der udvendigt er dækket af en pinacoderm. Når de gunstige forhold vender tilbage, vokser disse reduktionslegemer til helt nye svampe.
(iii) knopdannelse:
I svampe finder knopdannelse sted på forskellige måder.
(a) Eksogen knopdannelse:
En svamp danner udvendige knopper vegetativt ved basis af grene og danner derved en gruppe af individer. Til sidst forsnævrer knopperne sig fra forældrene, og hver enkelt danner en ny svamp.
(b) Endogen knopdannelse:
Skønnede formeringslegemer, kaldet gemmuler, dannes indvendigt hos alle ferskvandssvampe og nogle havsvampe.
Archeocytter samles i grupper i mesogloea, de bliver derefter flerkernede, de bliver også fyldt med proteiner som reservefødemateriale, der tilføres archeocytter af særlige ammeceller trophocytter. Nogle amøbocytter omslutter denne masse af fødefyldte archeocytter og udskiller en hård dobbeltlaget skal, skallen har et lille udløb eller en mikropyle.
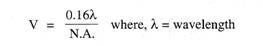
Dernæst udskiller nogle scleroblaster spiculer, som er placeret radialt mellem to lag af skallen, nogle spiculer rager ud uden for skallen, spiculerne i Spongilla er monaxonspiculer, men hos andre er de amfidisker. Amphidiskerne er lige stænger med tornede sider og en ring af kroge i hver ende.
Sådan dannes en gemmule, hvorefter de omgivende amøbocytter, scleroblaster og trophocytter afgår. Gemmuler dannes i stort antal om efteråret, hvorefter svampen opløses, de forbliver i svampens rester eller bliver frie, under alle omstændigheder falder de ned på bunden. Gemmules kan tåle ugunstige forhold, men de klækkes, når foråret kommer.
Ved udklækningen strømmer arkeocytterne ud af mikropylen, hvorefter disse flerkernede arkeocytter deler sig og danner ukernede arkeocytter og små celler kaldet histoblaster.
Histoblasterne danner ved differentiering og omlægning epidermisporocytter, choanocytter og indre endodermforing; modificerede archeocytter fra scleroblast, der udskiller spicula. Omkring en uges tid efter udklækningen omgiver en ung svamp den tomme gemmelskal. De ukernede archeocytter forbliver embryonale.

2. Seksuel forplantning:
Svampe har ingen kønsorganer, men amoebocytter danner kønsceller i mesenchymet ; først produceres æg og senere sædceller, derfor er svampen protogyn, hvor krydsbefrugtning finder sted. Amøbocytterne bliver fyldt med føde og bliver store, de bliver runde og danner æg.
Andre amøboocytter deler sig for at producere et stort antal sædceller, en sædcelle har et ovalt hoved og en lang tilspidset hale.
Nogle arbejdere hævder, at der dannes kønsceller fra archeocytter eller endog fra choanocytter. Æg fra en svamp befrugtes af sædceller fra en anden svamp, så der dannes zygoter. Ved krydsbefrugtning går sædcellen sandsynligvis ind i en choanocyt, som fusionerer med ægget og frigør sædcellen, som derefter fusionerer med ægget. Zygoten udskiller et hylster kaldet yngelkapsel, som omslutter zygoten.

Svampes udvikling:
Zygoten gennemgår en holoblastisk, men ulige spaltning; ved den holoblastiske deling er zygoten fuldstændig segmenteret.
De første tre delinger er vertikale, som danner otte pyramideceller, derefter danner en horisontal deling otte små øvre celler ved den animalske pol og otte store nedre celler ved den vegetative pol. De øverste små celler deler sig hurtigt, bliver klare og får flageller, mens de nederste celler deler sig langsomt og bliver granulerede.
Dette danner en blastula, som har et blastocoel hulrum indvendigt. Blastulaen kaldes en amfiblastula efter dannelsen af flageller. Hidtil har udviklingen fundet sted i svampens krop, nu finder amfiblastulaen vej ind i udstrømningskanalerne og forlader moderdyret gennem osculum.
Aphiblastulaen svømmer frit omkring i nogle timer, derefter invaginerer de øverste flagellede celler ind i blastocoelen, og de bliver overvokset af de nederste granulære celler; dette danner en gastrula, der ligner en kop med et ydre lag af granulære celler kaldet ektoderm og et indre lag af flagellede celler kaldet endoderm, den har en stor åbning, blastoporen.
Dette embryo hæfter sig ved en fast genstand ved sin blastoporale ende, og det begynder at vokse. Både ektoderm- og endodermlagene udskiller mesogloea og dets amøbocytter. Ifølge nogle udskilles mesogloea kun af choanocytter (endoderm). Væggen bliver tykkere og foldes og danner kanaler, og perforeringer danner ostia og osculum.

Kimlagene hos svampe svarer ikke til ektoderm og endoderm hos Metazoa, fordi svampens ydre ektoderm er blevet dannet af de nederste granulære celler fra den vegetative pol, og endoderm er dannet af de øverste flagellerede celler fra den animalske pol. I Metazoa bliver cellerne fra dyrepolen til ektoderm, og cellerne fra plantepolen danner endoderm.
Svampernes animalske natur:
I lang tid blev svampe ikke anset for at være dyr, netop på grund af deres ikke-animalske udseende; de er siddende og har ikke evnen til at fange føde eller slippe af med deres affaldsstoffer, og de udviser mere eller mindre ingen reaktion på stimuli.
Det var imidlertid Robert Grant (1836), der anerkendte svampenes dyriske karakter ved at foreslå, at vandstrømmen løber gennem en bestemt vej i svampekroppen; den trænger ind i svampekroppen gennem små porer, kaldet ostier, der er spredt ud over hele kroppens overflade, og passerer ud gennem større huller, kaldet oscula.
Sponges holozoiske ernæringsmåde; de ernærer sig af faste fødepartikler, der kommer med vandstrømmen, uden cellulosecellevæg, og tilstedeværelsen af frit svømmende larver med cilier i deres livscyklus, der ligner andre havdyrs, tyder klart på, at svampene er dyr og intet andet.
Affiniteter hos svampe:
Der har været en stor uenighed om svampenes natur og slægtskab, lige siden de blev opdaget. Aristoteles (384-322 f.Kr.) var den første til at anerkende dem som dyr. Men biologerne efter Aristoteles troede i århundreder, at de var planter på grund af deres fastsiddende vækstform og ufølsomme natur.
Ellis (1765) påpegede, at en kontinuerlig vandstrøm går ind i og ud af svampekroppen; det blev yderligere bekræftet af Robert Grant (1836). Linné, Lamarck og Cuvier placerede dem imidlertid sammen med coelenteraterater i Zoophyta. Robert Grant (1836) etablerede endelig en separat gruppe Porifera for svampene. De viser således slægtskab med protozoer såvel som med metazoer.
Affiniteter med protozoer:
Lighedspunkter:
Lighedspunkter:
Svampe udviser ligheder med koloniale protozoer ved at have følgende kendetegn:
Svampe viser ligheder med koloniale protozoer:
1. Fravær af fordøjelseshule og tilstedeværelse af intracellulær fordøjelse.
2. Tilstedeværelsen af kraveceller og amøboide celler som hos koloniale flagellater.
3. Produktion af skelet af en enkelt celle eller en gruppe af celler.
4. Cellerne i svampekroppen er indbyrdes afhængige i deres funktion.
5. En proces kendt som inversion forekommer i amfiblastula-larver som dem af Volvox. De flagellerede svampelarver betragtes som rester af en eller anden flagelleret kolonial protozoisk forfader.
Differencer:
Svampe adskiller sig fra protozoer ved at have følgende kendetegn:
Svampe adskiller sig fra protozoer ved at have følgende kendetegn:
1. Kanalsystemet.
2. Det karakteristiske skelet.
3. Udviklingen af et flercelligt legeme fra et befrugtet æg ved spaltning; og i denne egenskab bliver svampene mere komplekse end koloniale protozoer og ligner Metazoa’erne. Derfor står dens protozoiske slægtskab ikke længere.
Affiniteter med Metazoa:
Lighedspunkter:
Lighedspunkter:
I blandt Metazoa udviser coelenteraterne visse ligheder med svampene i følgende træk:
1. Begge har en stillesiddende levevis.
2. Begge er diploblastiske og acoelomate. Der findes et ikke-cellulært mesoglelag mellem ektoderm og endoderm. Mesoderm er fraværende i begge tilfælde.
3. Svampesvampes spongocoel, der åbner sig ud gennem terminal osculum, kan sammenlignes med det gastrovaskulære hulrum, der åbner sig udadtil gennem terminal hypos tome hos coelenteraterater.
4. Der sker aseksuel formering i begge tilfælde, og kolonierne dannes ved knopskydning.
5. Parenchymula-larven hos svampe er sammenlignelig med planulaen hos coelenteraterater.
forskelle:
Svampene adskiller sig fra Metazoa ved at have følgende kendetegn:
1. Hos svampene er cellerne forholdsvis mindre specialiserede og indbyrdes afhængige end cellerne hos Metazoa.
2. Svampe besidder choanocytter, som ikke findes hos nogen Metazoa.
3. Svampe er opbygget på celleplan, mens Metazoa er opbygget på vævs- og organplan.
4. Svampene er diploblastiske, mens Metazoa (undtagen Coelenterata) er triploblastiske.
5. Svampene besidder ikke en forreste ende eller et hoved som hos Metazoa. Osculumet fungerer kun som udåndingsåbning.
5. Svampenes legeme er perforeret af talrige små porer, og de besidder et enestående system af kanaler i deres legeme; alle disse findes aldrig hos Metazoa.
6. Svampe har ingen nervevæv til at koordinere kroppens aktiviteter.
7. Der er fuldstændig mangel på fordøjelseshule hos svampene i modsætning til Metazoa.
8. Svampe har et større antal fedtstoffer og fedtsyrer, som har en højere molekylvægt end dem, der findes i Metazoa.
9. I udviklingen af svampe bliver de flagellerede øvre celler i embryoet, der befinder sig ved den animalske pol, til endoderm, og de granulære celler i den vegetale pol bliver til ektoderm; hos ingen Metazoa forekommer en sådan omvending under udviklingen.
10. På grund af disse forskelle kan svampene ikke grupperes sammen med Metazoa, og der er derfor oprettet en særskilt stamme for svampene.
Så på trods af deres ligheder med protozoer og metazoer mener man, at svampene har afveget fra hovedlinjen i udviklingen af metazoer, og at de er et fylum i en blindgyde. I betragtning af dette er der blevet oprettet et særskilt underrige Parazoa, som omfatter svampene, og derfor har de en isoleret fylogenetisk position.
Svampes økonomiske betydning:
Svampene er af stor økonomisk betydning, både gavnlige og skadelige, for mennesket.
A. Nyttebringende svampe:
Svampene er gavnlige for mennesket og andre dyr på følgende måde:
1. Som føde:
Svampe bliver sjældent taget som føde af andre dyr på grund af deres dårlige smag, lugt og skarpe pigge. Der findes dog krebsdyr, der lever et parasitisk liv på dem, og nogle bløddyr som f.eks. nøgensnegle er afhængige af dem i deres kost.
2. Som kommensaler:
Svampe tjener som beskyttende huse for adskillige dyr som krebsdyr, orme, bløddyr, småfisk osv., fordi deres fjender ikke kan æde svampene. Ud over beskyttelsen får de dyr, der lever inde i svampekroppen, et rigt fødeudbud fra det vand, der cirkulerer gennem dem. Disse dyr forårsager ikke nogen tilsyneladende skade på svampene, de betragtes som kommensale dyr.
3. Andre anvendelsesmuligheder:
Det tørrede, fibrøse skelet af mange svampe som Spongia, Hippospongia og Euspongia bruges til badning, polering, vask af biler, vægge, møbler og gulvvask osv. Skelettet af nogle svampe som Euplectella er af stor kommerciel værdi og anvendes som dekorationsgenstande.
B. Skadelige svampe:
Betragtes kun nogle få svampe som skadelige. De kan forårsage visse fastsiddende dyrs død ved at vokse over dem og afskære deres føde- og iltforsyning. De kedelige svampe, som Cliona, sætter sig fast på skallerne af østers, muslinger og havtasker osv. Den borer sig ind i disse dyrs skaller og ødelægger dem fuldstændigt.
De borende svampe forårsager også stor skade på østersbanker. De borende svampe ødelægger også klipper ved at trænge ind i dem og bryde dem i stykker.
