Viruksen rakenne
Virukset eivät ole kasveja, eläimiä tai bakteereja, mutta ne ovat elävien valtakuntien keskeisiä loisia. Vaikka ne saattavat vaikuttaa eläviltä organismeilta ihmeellisten lisääntymiskykyjensä vuoksi, virukset eivät ole eläviä organismeja sanan varsinaisessa merkityksessä.
Ilman isäntäsolua virukset eivät voi suorittaa elämää ylläpitäviä toimintojaan tai lisääntyä. Ne eivät voi syntetisoida proteiineja, koska niiltä puuttuvat ribosomit ja niiden on käytettävä isäntäsolujensa ribosomeja kääntääkseen viruksen lähetti- RNA:n virusproteiineiksi. Virukset eivät voi tuottaa tai varastoida energiaa adenosiinitrifosfaatin (ATP) muodossa, vaan niiden on saatava energiansa ja kaikki muut aineenvaihduntatoimintonsa isäntäsolusta. Ne myös loisivat solusta perusrakennusaineita, kuten aminohappoja, nukleotideja ja lipidejä (rasvoja). Vaikka virusten on arveltu olevan eräänlainen protolife, niiden kyvyttömyys selviytyä ilman eläviä organismeja tekee erittäin epätodennäköiseksi, että ne olisivat edeltäneet soluelämää maapallon varhaisessa evoluutiossa. Jotkut tiedemiehet arvelevat, että virukset ovat alkaneet geneettisen koodin roistopätkistä, jotka ovat sopeutuneet loismaiseen elämään.
Kaikki virukset sisältävät nukleiinihappoa, joko DNA:ta tai RNA:ta (mutta eivät molempia), ja proteiinikuoren, joka ympäröi nukleiinihappoa. Joitakin viruksia ympäröi myös rasva- ja proteiinimolekyyleistä koostuva kuori. Tarttuvassa muodossaan, solun ulkopuolella, virushiukkasta kutsutaan virioniksi. Kukin virioni sisältää vähintään yhden ainutlaatuisen proteiinin, joka syntetisoidaan sen nukleiinihapon tiettyjen geenien avulla. Viroidit (tarkoittaa ”viruksen kaltaisia”) ovat tauteja aiheuttavia organismeja, jotka sisältävät vain nukleiinihappoa eikä niillä ole rakenneproteiineja. Muut viruksen kaltaiset hiukkaset, joita kutsutaan prioneiksi, koostuvat pääasiassa proteiinista, joka on tiiviisti integroitunut pieneen nukleiinihappomolekyyliin.
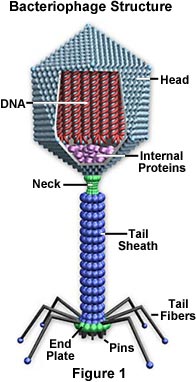
Virukset luokitellaan yleensä sen mukaan, mitä organismeja ne infektoivat, eläimiä, kasveja tai bakteereja. Koska virukset eivät pysty läpäisemään kasvien soluseinämiä, lähes kaikki kasvivirukset tarttuvat hyönteisten tai muiden kasveja syövien organismien välityksellä. Tietyt bakteerivirukset, kuten T4-bakteriofagi, ovat kehittäneet monimutkaisen tartuntaprosessin. Viruksella on ”häntä”, jonka se kiinnittää bakteerin pintaan proteiinipitoisten ”tappien” avulla. Häntä supistuu ja häntätulppa tunkeutuu soluseinään ja sen alla olevaan kalvoon, jolloin viruksen nukleiinihapot ruiskutetaan soluun. Virukset luokitellaan edelleen perheisiin ja sukuihin kolmen rakenteellisen seikan perusteella: 1) niiden nukleiinihapon tyyppi ja koko, 2) kapsidin koko ja muoto ja 3) se, onko niillä nukleokapsidia (kapsidin ympäröimää nukleiinihappoa) ympäröivä lipidikuori.
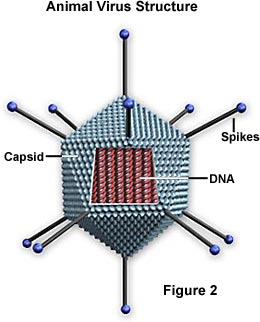
Virusten joukossa on pääasiassa kahdenlaisia muotoja: sauvoja eli filamentteja ja palloja. Sauvamuoto johtuu kapsidin muodostavien nukleiinihapon ja proteiinien alayksiköiden lineaarisesta asettelusta. Pallomainen muoto on itse asiassa 20-sivuinen monikulmio (ikosaedri).
Virusten luonne ymmärrettiin vasta 1900-luvulla, mutta niiden vaikutuksia oli havaittu vuosisatojen ajan. Brittiläinen lääkäri Edward Jenner löysi jopa rokottamisen periaatteen 1700-luvun lopulla havaittuaan, että lievän lehmärokko-taudin sairastaneet ihmiset olivat yleensä immuuneja tappavampaa isorokkoa vastaan. 1800-luvun lopulla tiedemiehet tiesivät, että jokin taudinaiheuttaja aiheutti tupakkakasvien taudin, mutta se ei kasvanut keinotekoisessa elatusaineessa (kuten bakteerit) ja oli liian pieni valomikroskoopilla havaittavaksi. Elävän soluviljelyn ja mikroskoopin kehittyminen 1900-luvulla antoi tutkijoille lopulta mahdollisuuden tunnistaa virukset. Genetiikan edistysaskeleet paransivat tunnistusprosessia dramaattisesti.
-
Kapsidi – Kapsidi on valkuaiskuori, joka ympäröi nukleiinihappoa; koteloidun nukleiinihapon kanssa sitä kutsutaan nukleokapsidiksi. Tämä kuori koostuu proteiinista, joka on järjestäytynyt kapsomeereiksi kutsuttuihin alayksiköihin. Ne ovat tiiviisti yhteydessä nukleiinihappoon ja heijastavat sen konfiguraatiota, joka on joko sauvanmuotoinen kierre tai monikulmion muotoinen pallo. Kapsidilla on kolme tehtävää: 1) se suojaa nukleiinihappoa entsyymien pilkkomiselta, 2) sen pinnalla on erityisiä kohtia, joiden avulla virioni voi kiinnittyä isäntäsoluun, ja 3) se tarjoaa proteiineja, joiden avulla virioni voi tunkeutua isäntäsolun kalvon läpi ja joissakin tapauksissa ruiskuttaa tartunnan aiheuttavan nukleiinihapon solun sytoplasmaan. Oikeissa olosuhteissa viruksen RNA proteiinimolekyylien nestemäisessä suspensiossa kokoaa kapsidin itsestään toimivaksi ja tarttuvaksi virukseksi.
-
Kuori – Monilla virustyypeillä on nukleokapsidia ympäröivä glykoproteiinikuori. Kuori koostuu kahdesta lipidikerroksesta, joiden välissä on proteiinimolekyylejä (lipoproteiinikaksoiskerros), ja se voi sisältää sekä isäntäsolun kalvosta peräisin olevaa materiaalia että virusperäistä materiaalia. Virus saa lipidimolekyylit solukalvolta viruksen budding-prosessin aikana. Virus kuitenkin korvaa solukalvon proteiinit omilla proteiineillaan, jolloin syntyy hybridirakenne, joka koostuu solusta peräisin olevista lipideistä ja viruksesta peräisin olevista proteiineista. Monet virukset kehittävät kuoriinsa myös glykoproteiinista valmistettuja piikkejä, jotka auttavat niitä kiinnittymään tietyille solupinnoille.
-
Nukleiinihappo – Aivan kuten soluissa, jokaisen viruksen nukleiinihappo koodaa geneettisen tiedon kaikkien proteiinien synteesiä varten. Vaikka kaksijuosteinen DNA vastaa tästä prokaryoottisissa ja eukaryoottisissa soluissa, vain muutamat virusryhmät käyttävät DNA:ta. Useimmat virukset säilyttävät kaiken geneettisen informaationsa yksijuosteisella RNA:lla. RNA-pohjaisia viruksia on kahdenlaisia. Useimmissa genomista RNA:ta kutsutaan plus-juosteeksi, koska se toimii lähetti- RNA:na viruksen proteiinin suoraa synteesiä (translaatiota) varten. Muutamilla on kuitenkin negatiivisia RNA-juosteita. Näissä tapauksissa virionissa on entsyymi, jota kutsutaan RNA-riippuvaiseksi RNA-polymeraasiksi (transkriptaasiksi), jonka on ensin katalysoitava komplementaarisen lähetti- RNA:n tuottaminen virion genomisesta RNA:sta ennen kuin viruksen proteiinisynteesi voi tapahtua.
Influenssa (influenssa) -virus – Influenssa eli ”influenssa” on flunssan ohella ehkä maailman tunnetuin hengitystieinfektio. Pelkästään Yhdysvalloissa noin 25-50 miljoonaa ihmistä sairastuu influenssaan vuosittain. Influenssan oireet muistuttavat flunssan oireita, mutta ovat yleensä vakavampia. Kuume, päänsärky, väsymys, lihasheikkous ja -kipu, kurkkukipu, kuiva yskä ja vuotava tai tukkoinen nenä ovat yleisiä ja voivat kehittyä nopeasti. Influenssaan liittyviä ruoansulatuskanavan oireita esiintyy joskus lapsilla, mutta useimmilla aikuisilla ripulina, pahoinvointina ja oksenteluna ilmenevät sairaudet eivät johdu influenssaviruksesta, vaikka niitä kutsutaankin usein virheellisesti ”vatsaflunssaksi”. Influenssan yhteydessä voi esiintyä myös useita komplikaatioita, kuten keuhkoputkentulehduksen ja keuhkokuumeen puhkeaminen, ja ne ovat erityisen yleisiä iäkkäillä, pienillä lapsilla ja kaikilla, joiden immuunijärjestelmä on heikentynyt.
Ihmisen immuunipuutosvirus (HIV) – HIV:n aiheuttavan viruksen eristivät ensimmäisen kerran vuonna 1983 yhdysvaltalainen Robert Gallo ja ranskalainen tutkija Luc Montagnier. Sen jälkeen on tehty valtava määrä AIDSin aiheuttajaan keskittyvää tutkimusta, ja paljon on opittu viruksen rakenteesta ja sen tyypillisestä toimintatavasta. HIV kuuluu ryhmään epätyypillisiä viruksia, joita kutsutaan retroviruksiksi ja jotka säilyttävät geneettisen informaationsa ribonukleiinihapon (RNA) muodossa. Käänteiseksi transkriptaasiksi kutsutun entsyymin avulla HIV ja muut retrovirukset pystyvät tuottamaan deoksiribonukleiinihappoa (DNA) RNA:sta, kun taas useimmat solut suorittavat päinvastaisen prosessin, DNA:n geneettisen materiaalin transkriboimisen RNA:ksi. Entsyymin toiminnan ansiosta HIV:n geneettinen informaatio integroituu pysyvästi isäntäsolun genomiin (kromosomeihin).
TAKAISIN SELLIRAKENTEEN KOTISIVULLE
Kysymyksiä tai kommentteja? Lähetä meille sähköpostia.
© 1995-2021 by Michael W. Davidson and The Florida State University. Kaikki oikeudet pidätetään. Kuvia, grafiikkaa, ohjelmistoja, skriptejä tai appletteja ei saa jäljentää tai käyttää millään tavalla ilman tekijänoikeuksien haltijoiden lupaa. Tämän verkkosivuston käyttö tarkoittaa, että hyväksyt kaikki omistajien asettamat oikeudelliset ehdot.
Tämän verkkosivuston ylläpitäjänä toimii
Graphics & Web Programming Team
yhteistyössä Optical Microscopy at the
National High Magnetic Field Laboratory:n
kansallisen korkean magneettikentän laboratorion kanssa.
Viimeisin muutos:
Access Count Since October 1, 2000: 1951931
Microscopes provided by:
![]()
![]()