B -globiinin geeniperhe
Ihmisen genomi koostuu noin 3 miljardista emäsparista. Viime aikoihin asti tutkijat olivat ennustaneet, että tämä suuri määrä DNA:ta koodaisi lähes 100 000 erilaista geeniä; nyt kun koko ihmisen perimän sekvensointi on kuitenkin saatu lähes valmiiksi, tämä luku on pudonnut vain 30 000 geeniin. Itse asiassa genomin tarkka analyysi on osoittanut, että reilusti yli 90 prosenttia genomista koostuu ei-toimivasta DNA:sta.
Jäljelle jäävästä toiminnallisesta DNA:sta 25-50 prosenttia proteiineja koodaavista geeneistä esiintyy vain kerran haploidissa genomissa, ja niitä kutsutaan yksinäisiksi geeneiksi. Usein tiettyä geeniä ympäröivä DNA sisältää sekvenssejä, jotka ovat läheisiä mutta eivät identtisiä kopioita geenistä. Näiden moninkertaisten kopioiden uskotaan olevan seurausta duplikaatiosta ja divergenssistä. Tämä on prosessi, jossa yksittäinen geeni ensin monistuu ja joutuu sitten alttiiksi valikoivalle paineelle muuntua geeniksi, joka on sekvenssiltään samankaltainen mutta ei identtinen esi-isänsä geenin kanssa. Yksi esimerkki tästä prosessista on beetaglobiinigeeniperheen kehitys. Kun eri lajien globiiniperheen geenit oli sekvensoitu, kehitettiin evoluutiopuu, jossa ennustettiin globiiniperheen geenien kehitystä. Alla on esitetty ehdotettu puu, joka havainnollistaa geenien erilaistumista kasvien leghemoglobiinista eläinten hemoglobiiniin ja myoglobiiniin. Jokaisessa haarautumiskohdassa geeni on monistunut ja mutatoitunut uudeksi, mutta samanlaiseksi geeniksi. Esimerkiksi jossain vaiheessa historiaa yhteinen esi-isägeeni koki ensin monistumisen. Syntyneet kaksi kopiota mutatoituivat sitten eri tavoin; toinen muodosti jalkahemoglobiinin toinen hyönteisten hemoglobiinin.
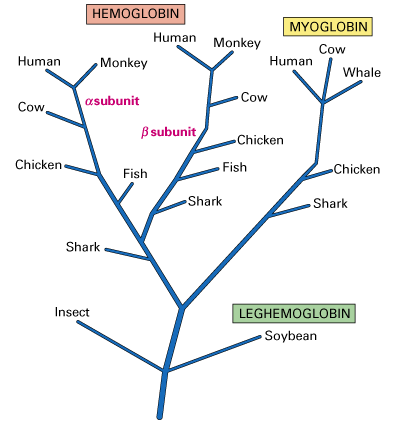
Kuvio 1: Kaavio globiinisukupuusta (Lodish et.al.,2000)
Sarjaa monistuneista geeneistä, jotka koodaavat proteiineja, joilla on samanlainen mutta ei-identtinen aminohapposekvenssi, sanotaan geenisukupuuksi (Lodish et.al., 2000). On tunnistettu lukuisia erilaisia geeniperheitä, joilla on erilaisia tehtäviä. Esimerkiksi toimivan hemoglobiinimolekyylin muodostaminen edellyttää kahden tällaisen geeniperheen tuotteiden käyttöä yhdistämällä kaksi b -globiiniperheen polypeptidiä kahteen a -globiiniperheen polypeptidiin ja neljään pieneen hemiryhmään. Kromosomissa 11 sijaitseva beetaglobiinigeeniperhe on esitetty alla olevassa kuvassa, ja se koostuu viidestä toiminnallisesta geenistä (siniset laatikot) ja kahdesta pseudogeenistä (vinoviivat).
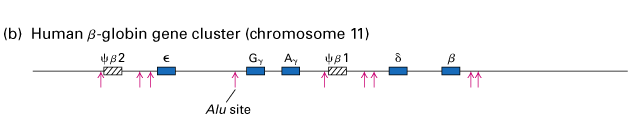
Kuva 2: Kromosomissa 11 sijaitseva beetaglobiinigeeniperhe (Lodish ym., 2000)
Kaikkien näiden erilaisten geenien koodaamien hemoglobiinien tehtävänä on kuljettamaan happea veressä; kullakin geeneillä on kuitenkin erityisiä toiminnallisia vaihteluita. Esimerkiksi epsilon-globiinigeeni ilmentyy normaalisti alkion keltarauhasessa, kun taas Ag- ja Gg-geenit ilmentyvät vain sikiön kehityksen aikana. Näillä hemoglobiiniproteiineilla on suurempi hapen sitoutumisaffiniteetti kuin b- ja d-geenien koodaamilla aikuisten hemoglobiineilla. Tämän lisääntyneen sitoutumisen ansiosta sikiö voi menestyksekkäästi ottaa happea verestä kilpailematta äidin kanssa. Aikuisten hemoglobiinin happiaffiniteetti on alhaisempi, mikä mahdollistaa hapen paremman vapautumisen kudoksiin, erityisesti lihaksiin.
Globiinigeeniperheen kaksi aluetta sisältävät ei-toiminnallisia sekvenssejä, joita kutsutaan pseudogeeneiksi (vinoviivoitetut laatikot). Nämä geenit ovat samankaltaisia kuin niiden funktionaaliset globiinigeenit, mutta niitä ei enää transkriboida mRNA:ksi niiden evoluution aikana tapahtuneiden sekvenssimuutosten vuoksi.
Jopa pienikin muutos jossakin hemoglobiinimolekyylin alayksikköä koodaavassa geenissä voi johtaa katastrofaalisiin seurauksiin. Tässä laboratoriossa keskitytään kahteen sairauteen, jotka johtuvat erilaisista mutaatioista beetaglobiinigeenissä.
Klikkaa alla olevaa linkkiä saadaksesi lisätietoa sirppisoluanemiasta.