ADVERTISSEMENTS:
Dans cet article, nous allons discuter des éponges:- 1. Histoire des éponges 2. Définition des éponges 3. Origine 4. caractères généraux 5. Classification 6. Organisation générale 7. Système de canaux 8. Squelette 9. Reproduction 10. Le développement 11. La nature animale 12. Affinités 13. Importance économique.
Contenu :
- Histoire des spongiaires
- Définition des spongiaires
- Origine des spongiaires
- Caractères généraux des spongiaires
- Classification des spongiaires
- Organisation générale des spongiaires
- Système canalaire chez les… Sponges
- Squelette des éponges
- Reproduction des éponges
- Développement des éponges
- Nature animale des éponges
- Affinités des éponges
- Importance économique des éponges
Histoire des éponges :
Le nom Porifera (L., porus = pore ; ferro = porter) vient de (1836). La nature des éponges a été débattue jusqu’à une bonne partie du XIXe siècle, bien que la preuve de leur nature animale ait été donnée en 1765 par celui qui a vu les courants d’eau et les mouvements de l’oscula. En conséquence, Linnaeus, Lamarck, et ont classé les éponges dans la catégorie des zoophytes ou des polypes et les ont considérées comme alliées aux coelentérés anthozoaires.
Avertissements:
Bien que (1816) ait séparé les éponges dans un groupe Spongiaria allié aux Protozoaires. La morphologie et la physiologie des éponges ont d’abord été convenablement comprises par qui a créé en 1836 le nom Porifera pour le groupe par lequel il est maintenant généralement connu, iuxle (1875) et Sollas (1884) ont proposé la séparation complète des éponges des autres Metazoa sur la base de nombreuses particularités.
Les éponges sont maintenant reconnues comme constituant une branche distincte et isolée des Métazoaires nommée Parazoa, d’après Sollas.

Les Porifères comprennent les éponges qui sont les plus primitifs des animaux multicellulaires, ce sont des animaux sessiles, semblables à des plantes, ils sont fixés à quelque roche solide ou coquille submergée et sont incapables de tout mouvement. Les Porifères sont exclusivement marins, à l’exception d’une seule famille d’espèces d’eau douce.
Avertissements:
Leur forme peut être cylindrique, ramifiée, en vase ou globulaire, certaines sont de couleur terne mais la plupart sont de couleur vive, elles ont une couleur rouge, orange, violette, verte ou jaune. Le corps est perforé par des pores et des canaux mais il n’y a pas d’organes, comme la bouche ou le système nerveux. Bien que les éponges soient des animaux multicellulaires, leurs cellules ne forment pas de tissus organisés. Elles possèdent généralement un endosquelette constitué de spicules séparés.
La digestion a lieu à l’intérieur des cellules. En raison de leur endosquelette et de leurs ferments odieux, elles ne sont généralement pas consommées par les animaux. Les éponges sont cultivées à des fins commerciales.
On connaît actuellement environ 10 000 espèces d’éponges et l’embranchement est divisé en trois classes, à savoir, Calcarea ou Calcispongiae, Hexactinellida ou Hyalospongiae, et Demospongiae et une douzaine d’ordres principalement sur le type de squelette.
Définition des spongiaires :
Les Porifères peuvent être définis comme des « organismes multicellulaires asymétriques ou radialement symétriques avec un grade d’organisation cellulaire sans tissus et organes bien définis ; exclusivement aquatiques ; principalement marins, sédentaires, animaux solitaires ou coloniaux avec un corps perforé par des pores, des canaux et des chambres à travers lesquels l’eau circule ; avec une ou plusieurs cavités internes tapissées de choanocytes ; et avec un squelette caractéristique fait de spicules calcaires, de spicules siliceuses ou de fibres cornées de spongine ».
Origine des éponges :
Il existe une grande controverse concernant l’origine des Porifères. Cependant, ils semblent plus proches des Protozoaires bien qu’ils s’en distinguent en étant multicellulaires. Les Porifères sont probablement issus de protozoaires flagellés comme Proterospongia, un flagellé colonial. La colonie de Proterospongia possède des cellules collées et flagellées noyées dans une matrice gélatineuse comportant des cellules amiboïdes.
Ces cellules sont les choanocytes et les amibocytes des spongiaires. Même dans ce cas, on pense que l’origine des éponges est incertaine. De manière concluante, on peut dire que, les éponges ont divergé de la lignée évolutive des métazoaires à un stade très précoce et, par conséquent, occupent un statut distinct.
Caractères généraux des spongiaires :
1. Les Porifères sont tous aquatiques, principalement marins sauf une famille Spongillidae qui vit en eau douce.
2. Elles sont sessiles et sédentaires et poussent comme des plantes.
Avertissements :
3. La forme du corps est en vase ou en cylindre, asymétrique ou à symétrie radiale.
4 La surface du corps est perforée par de nombreux pores, les ostia par lesquels l’eau entre dans le corps et une ou plusieurs grandes ouvertures, les oscula par lesquels l’eau sort.
5. Corps multicellulaire constitué d’un ectoderme externe et d’un endoderme interne avec une couche intermédiaire de mésenchyme, donc des animaux diploblastiques.

6. L’espace intérieur du corps est soit creux, soit perméable par de nombreux canaux tapissés de choanocytes. L’espace intérieur du corps de l’éponge est appelé spongocoel.
Avertissements:
7. Squelette caractéristique constitué soit de fines fibres spongieuses flexibles, soit de spicules siliceux, soit de spicules calcaires.
8. Bouche absente, digestion intracellulaire ; organes excréteurs et respiratoires absents.
9. Les cellules nerveuses et sensorielles ne sont probablement pas différenciées.
PUBLICITÉS:
10. Les éponges sont monoïques ; reproduction à la fois par des méthodes asexuées et sexuées.
11. La reproduction asexuée se fait par les bourgeons et les gemmules.
12. Les éponges possèdent un grand pouvoir de régénération.
13. La reproduction sexuée se fait par des ovules et des spermatozoïdes ; la fécondation est interne mais la fécondation croisée se produit en règle générale.
PUBLICITÉS:
14. Clivage holoblastique, développement indirect par une larve ciliée à nage libre appelée amphiblastula ou parenchymula.
15. L’organisation des éponges a été regroupée en trois grands types, à savoir, le type ascon, le type sycon et le type leuconoïde en raison de la simplicité de certaines formes et de la complexité d’autres.
Classification des spongiaires :
La classification des Porifères est basée principalement sur les types de squelette qu’on y trouve. Ce phylum a été classé diversement mais la classification proposée par Hyman (1940) et Burton (1967) est d’une importance considérable. Cependant, la classification des Porifera suivie ici est basée sur Storer et Usinger (1971) qui semble être une forme modifiée de la classification de Hyman.
Classe I. Calcarea (L., Calx = Chaux) ou Calcispongiae :
(L., calx = chaux,Gr., spongos = éponge) :
1. Ils ont un squelette de spicules calcaires séparés qui sont monaxons ou tétraxons ; les spicules tétraxons perdent un rayon pour devenir triadiés.
ADVERTISSEMENTS :
2. Ils sont solitaires ou coloniaux ; forme du corps en vase ou cylindrique.
3. ils peuvent présenter une structure asconoïde, syconoïde ou leuconoïde.
4. Ce sont des éponges de couleur terne dont la taille est inférieure à 15 cm.
5. Elles sont présentes dans les eaux peu profondes de tous les océans.
Ordre 1. Homocoela :
1. Éponges asconoïdes à corps cylindrique à symétrie radiale.
Avertissements :
2. La paroi du corps est mince et non pliée, le spongocoel est tapissé de choanocytes.
Exemples :
Leucosolenia, Clathrina.
Ordre 2. Heterocoela :
1. Les éponges syconoïdes ou leuconoïdes ayant un corps en forme de vase.
2. La paroi du corps est épaisse et plissée, les choanocytes ne bordent que les canaux radiaux.
3. Le spongocoel est tapissé par des cellules endodermiques aplaties.
Exemples :
Sycon ou Scypha, Grantia.
Classe. II. Hexactinellida ou Hyalospongiae :
(Gr., hyalos = vitreux ; spongos = éponge) :
1. On les appelle éponges de verre ; le squelette est constitué de spicules siliceuses qui sont triaxones à six rayons. Chez certaines, les spicules sont soudées pour former un squelette en treillis.
2. Il n’y a pas d’épithélium épidermique.
3. Les choanocytes tapissent des chambres en forme de doigts.
4. Ils sont cylindriques ou en forme d’entonnoir et se trouvent dans les mers tropicales profondes, ils peuvent atteindre un mètre.
Ordre 1. Hexasterophora :
1. Les spicules sont hexasters, c’est-à-dire en forme d’étoile.
2. Les canaux radiaux ou les chambres flagellées sont simples.
3. Ils ne sont pas attachés par des touffes de racines mais communément attachés à un objet dur.
Exemples :
Euplectella, Farnera.
Ordre 2. Amphidiscophora :
1. Les spicules sont des amphidiscs. Pas d’hexasters.
2. Ils sont fixés au substrat par des touffes de racines.
Exemples :
Hyalonema, Pheronema.
Classe III. Demospongiae (Gr., dermos = cadre ; spongos = éponge) :
1. Elle contient le plus grand nombre d’espèces d’éponges. De grande taille, solitaires ou coloniales.
2. le squelette peut être constitué de fibres spongieuses ou de fibres spongieuses avec des spicules siliceux ou il peut ne pas y avoir de squelette.
3 Les spicules ne sont jamais à six rayons, ils sont monaxons ou tétraxons et sont différenciés en grands mégasclères et petits microsclères.
4. La forme du corps est irrégulière et le système de canaux est de type leucon.
5. Généralement marins, quelques formes d’eau douce.
Sous-classe I. Tetractinellida :
1. Les éponges sont pour la plupart solides et simples, arrondies en forme de coussin aplati, généralement sans branches.
2. Squelette composé principalement de spicules siliceux tétraxons mais absent dans l’ordre des Myxospongida.
3. Le système de canaux est de type leuconoïde. Forme d’eau peu profonde.
Ordre 1. Myxospongida :
1. Structure simple.
2. Squelette absent.
Exemples :
Oscarella, Halisarca.
Ordre 2. Carnosa :
1. Structure simple.
2. Les spicules ne sont pas différenciés en mégascléres et microscléres.
3. Des asters peuvent être présents.
Exemple :
Plakina.
Ordre 3. Choristide :
1. Les spicules sont différenciés en mégascléres et microscléres.
Exemples :
Geodia, Thenea.
Sous-classe II. Monaxonida :
1. Les monaxonides se présentent sous diverses formes allant de la masse arrondie au type ramifié ou allongé ou pédonculé avec entonnoir ou en éventail.
2 Le squelette est constitué de spicules de monaxons avec ou sans spongine.
3. Les spicules se distinguent en mégasclères et microsclères.
4. On les trouve en abondance dans le monde entier.
5. Des formes d’eau peu profonde et d’eau profonde.
Ordre 1. Hadromerida :
1. Mégasclères monaxons sous forme de tylostyles.
2. les microsclères lorsqu’ils sont présents sous forme d’asters.
3. Les fibres de spongine sont absentes.
Exemples :
Cliona, Tethya.
Ordre 2. Halichondrida :
1. Les mégasclères monaxons sont souvent de deux types, à savoir les monactines et les diactines.
2. les microsclères sont absents. Les fibres de spongine sont présentes mais peu abondantes.
Exemple :
Halichondria.
Ordre 3. Poecilosclerida :
1. Les mégasclères monaxons sont de deux types, un type dans l’ectoderme et un autre type dans la couche des choanocytes.
2. Les mégasclères sont typiquement des chélas, des sigmas et des toxas.
Exemple :
Cladorhiza.
Ordre 4. Haplosclerida :
1. Les mégasclères monaxons sont d’un seul type, à savoir, diactinaux.
2. Les microsclères sont absents.
3. Les fibres spongieuses sont généralement présentes.
Exemples :
Chalina, Pachychalina, Spongilla
Sous-classe III. Keratosa :
1. Le corps est arrondi et massif avec un certain nombre d’oscula ostensibles.
2. Squelette composé d’un réseau de fibres spongieuses uniquement.
3. Les spicules siliceux sont absents.
4. Elles sont également connues sous le nom d’éponges cornées que l’on trouve dans les eaux peu profondes et chaudes des régions tropicales et subtropicales.
Exemples :
Euspongia, Hippospongia.

Organisation générale des spongiaires :
L’organisation générale des spongiaires varie considérablement. Les éponges sont cylindriques comme Leucosolenia, en forme de vase comme Scypha et Grantia, en forme d’arbre (par exemple, Microciona), en forme de doigt (par exemple, Haliclona), en forme de feuille (par exemple, Phyllospongia), en forme de coussin comme Euspongia, en forme de corde (par exemple, Hyalonema), en forme de bol comme Pheronema, etc. Certaines éponges sont solitaires, tandis que d’autres sont coloniales.
Les éponges sont surtout des formes fixées ; on les trouve fixées à des pierres, des coquillages, des bâtons, des algues, etc… ; certaines sont des éponges ennuyeuses comme Cliona. Généralement, le corps de l’éponge est asymétrique mais quelques formes présentent une symétrie radiale. La taille varie de quelques mm à massive ayant 1 ou 2 mètres de diamètre.
La coloration du corps des éponges varie également beaucoup ; elles sont le plus souvent blanches ou grises mais des espèces de couleur jaune, brune, violette, orange, rouge et verte sont également signalées. La couleur verte de l’éponge est généralement due à la présence d’une algue symbiotique, la chlorelle Zoo, en leur sein.

Système de canaux chez les éponges:
Toutes les cavités du corps traversées par les courants d’eau, qui nourrissent l’éponge depuis son entrée par les pores jusqu’à sa sortie par l’oscule, sont collectivement appelées système de canaux. Chez Olynthus, le système de canaux est vu dans son type le plus simple.
Dans d’autres formes, il peut atteindre un haut degré de complexité, mais son évolution générale peut néanmoins être réduite à un simple processus de croissance de la part de l’Olynthus primitif, résultant en un repliement des parois et accompagné d’une restriction des cellules à collerette (choanocyte) à certaines régions.
Dans le processus graduel et continu de différenciation, on peut distinguer trois types distincts d’organisation qui reliés par de nombreuses transitions peuvent encore être considérés comme trois styles d’architecture, pour ainsi dire sous lesquels toutes les formes existantes peuvent être classées. Il y a habituellement trois types de système de canaux rencontrés chez les éponges, à savoir, le type asconoïde, le type syconoïde et le type leuconoïde.
(i) Type asconoïde :
Le type asconoïde du système de canaux est le plus simple de tous les types. Dans celui-ci, il y a un corps en forme de vase à symétrie radiale constitué d’une paroi mince enfermant une grande cavité centrale le spongocoel s’ouvrant au sommet par l’osculum rétréci.
La paroi est composée d’un épithélium externe et interne avec un mésenchyme entre les deux. L’épithélium externe ou dermique, ici appelé épiderme, est constitué d’une seule couche de cellules plates. L’épithélium interne, qui tapisse le spongocœle, est composé de choanocytes. Le mésenchyme contient des spicules squelettiques et plusieurs types d’amibocytes, le tout noyé dans une matrice gélatineuse.
La paroi de l’éponge asconoïde est perforée par de nombreuses ouvertures microscopiques appelées pores ou ostia incurrents qui s’étendent de la surface externe au spongocœle. Chaque pore est intracellulaire, c’est-à-dire qu’il s’agit d’un canal traversant une cellule tubulaire appelée porocyte.

Le courant d’eau impulsé par les flagelles des choanocytes passe par les pores incurrents dans le spongocoel et sort par l’oscule (eau de l’extérieur → pores incurrents → spongocoel → oscule → eau sortie) fournissant sur son passage nourriture et oxygène et emportant les déchets métaboliques. Le type asconoïde du système de canaux ne se trouve que chez quelques éponges, par exemple Olynthus, Leucosolenia.
Selon Hyman, les caractéristiques importantes de la structure asconoïde sont la paroi simple et l’épithélium de revêtement complet et continu des choanocytes, interrompu seulement par les extrémités intérieures des porocytes. Le type d’éponge asconoïde ressemble superficiellement à une gastrula typique.
2. le type syconoïde :
Le type syconoïde du système de canaux est le premier stade au-dessus du type asconoïde. Il est formé par la poussée vers l’extérieur de la paroi d’une éponge asconoïde à intervalles réguliers en projections en forme de doigts, appelées canaux radiaux.
Au début, ces canaux radiaux sont des projections libres et l’eau extérieure entoure toute leur longueur, car il n’y a pas de canaux incurrents définis. Mais chez la plupart des éponges syconoïdes, les parois des canaux radiaux fusionnent de manière à laisser entre elles des espaces tubulaires, les canaux incurrents, qui s’ouvrent sur l’extérieur entre les extrémités extérieures aveugles des canaux radiaux par des ouvertures appelées ostia dermiques ou pores dermiques.
Comme ces canaux incurrents représentent la surface externe originale de l’éponge asconoïde, ils sont nécessairement tapissés d’épiderme. Les canaux radiaux étant les excroissances du spongocoel originel sont nécessairement tapissés de choanocytes et sont, par conséquent, mieux appelés canaux flagellés.
L’intérieur de l’éponge syconoïde est creux et forme un grand spongocoel qui est tapissé par l’épithélium plat dérivé de l’épiderme. Les ouvertures des canaux radiaux dans le spongocoel sont appelées ostia internes. Les éponges syconoïdes conservent la forme de vase radial des asconoïdes et le spongocoel s’ouvre sur l’extérieur par l’unique oscule terminal.
La paroi entre les canaux incurrents et radiaux, est percée de nombreux pores minuscules appelés prosoples. Le courant d’eau dans les éponges syconoïdes prend le chemin suivant pores dermiques → canaux incurrents → prosopyles → canaux radiaux → ostia internes (apopyles) → spongocoeL → osculum → sortie.

Les éponges syconoïdes diffèrent du type asconoïde par deux particularités importantes :
(i) Dans les parois épaisses et plissées contenant alternativement des canaux incurrents et radiaux et
(ii) Dans la rupture de la couche de choanocytes, qui ne tapisse plus tout l’intérieur mais se limite à certaines chambres définies (canaux radiaux).

La structure syconoïde se présente en deux stades principaux. Le premier type illustré dans quelques-unes des éponges calcaires hétérocoeles, en particulier les membres du genre Sycon. Dans le deuxième stade, l’épiderme et le mésenchyme s’étendent sur la surface externe en formant un cortex mince ou épais contenant souvent des spicules corticaux spéciaux. L’épiderme devient percé de pores plus définis qui débouchent sur des canaux incurrents rétrécis.
3. type leuconoïde :
A la suite d’un autre processus de repliement de la couche de choanocyte et d’épaississement de la paroi du corps, le type leuconoïde du système de canaux se développe. La couche de choanocyte du canal radial du stade syconoïde s’évagine en de nombreuses petites chambres, et celles-ci peuvent répéter le processus, de sorte que des grappes de petites chambres flagellées arrondies ou ovales remplacent les chambres allongées du stade syconoïde.
Les choanocytes sont limités à ces chambres. Le mésenchyme remplit les espaces autour des chambres flagellées. Le spongocoel est généralement oblitéré et l’ensemble de l’éponge devient de structure irrégulière et de forme indéfinie. L’intérieur de l’éponge devient perméable par de nombreux canaux incurrents et ex-currents se rejoignent pour former de plus grands canaux ex-currents et des espaces qui mènent à l’oscula.
La surface est couverte d’épithélium épidermique et est percée de nombreux pores dermiques (ostia) et d’oscula.
Les pores dermiques débouchent sur des canaux incurrents qui se ramifient irrégulièrement dans le mésenchyme. Les canaux incurrents débouchent sur les petites chambres flagellées arrondies par des ouvertures encore appelées prosoples. Les chambres flagellées s’ouvrent par des ouvertures appelées apopyles dans des canaux ex-currents, et ceux-ci s’unissent pour former des tubes de plus en plus grands, dont les plus grands conduisent à l’oscula.

Les principales caractéristiques du système canalaire de type leuconoïde sont la limitation des choanocytes à de petites chambres, le grand développement du mésenchyme et la complexité des canaux incurrents et excurrents.
Le parcours du courant d’eau est ostia dermique → canaux incurrents → prosodus (si présent) → prosopyles → chambres flagellées → apopyles → aphodus (si présent) → canaux excurrents → canaux plus larges → oscula → sortie. Le système de canaux de type leucon est très efficace et la plupart des éponges sont construites sur le plan leuconoïde et elles atteignent une taille considérable.
Elles ont toujours une structure irrégulière mais l’écoulement du courant de l’eau est assez rapide et efficace. Le système de canaux de type leuconoïde présente de nombreuses variations mais trois stades d’évolution : eurypyle, aphode et diplode.
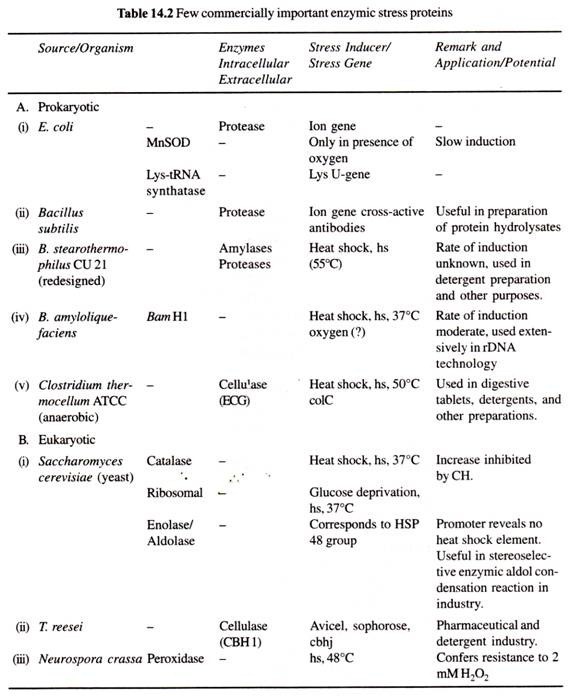
(a) Eurypyle :
Dans le système canalaire de type eurypyle leuconoïde, les chambres flagellées sont larges et en forme de dé à coudre, chacune débouchant directement dans le canal excurrent par une large ouverture appelée apopyle et recevant l’alimentation en eau directement du canal incurrent par le prosopyle.
Le courant d’eau emprunte le trajet suivant → pores ou ostia dermiques → espaces sous-cutanés → canaux incurrents → prosoples → chambres flagellées → apopyles -> canaux excurrents spongocoel → oscula → sortie. Ce type de système de canaux se retrouve chez Leucilla.
(b) Aphodale :
Dans le système de canaux de type leuconoïde aphodale, les chambres flagellées sont petites et arrondies. L’ouverture de chaque chambre flagellée dans le canal excurrent est dessinée dans un tube étroit, généralement pas de grande longueur, appelé aphodus. La relation entre les chambres flagellées et les canaux incurrents reste la même qu’auparavant.
Le trajet du courant d’eau est le suivant pores ou ostia dermiques → espace subdermique → canaux incurrents → prosoples → chambres flagellées → aphodus → canaux excurrents→ spongocoel → oscula → sortie. Ce type de système de canaux se retrouve chez Geodia et Stelleta.
(c) Diplodale :
Dans certains cas, il existe également un tube de courant étroit, le prosode entre le canal incurrent et les chambres flagellées, une telle condition est appelée diplodale. Ce type de système de canaux se retrouve chez Oscarella, Spongilla, etc.
Le courant d’eau prend le chemin suivant :
Pores ou ostia dermiques → espaces sous-cutanés → canaux incurrents → prosodus → chambres flagellées → aphodus → canaux excurrents → spongocoel → oscula → sortie.
4. Type Rhagon :
Dans les éponges calcaires, la structure leuconoïde peut être atteinte en passant par des stades asconoïdes et syconoïdes. Mais chez les Demospongiae, elle est dérivée d’un stade appelé rhagon qui, à son tour, résulte d’un réarrangement direct de la masse cellulaire interne.

Le type rhagon de l’éponge a une base large et il est de forme conique avec un seul oscule au sommet. La paroi basale est appelée l’hypophare qui est dépourvue de chambres flagellées. La paroi supérieure portant une rangée de petites chambres ovales flagellées est appelée spongophare.
Le spongocoel est bordé de chambres ovales flagellées s’ouvrant en son sein par de larges apopyles. Entre les chambres et l’épiderme se trouve une épaisseur considérable de mésenchyme traversé par des canaux incurrents et des espaces sous-dermiques. Les pores ou ostia dermiques s’ouvrent dans des espaces sous-dermiques qui s’étendent sous toute la surface du corps.
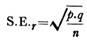
Des canaux incurrents ramifiés conduisent des espaces sous-dermiques dans de petites chambres flagellées qui ont été formées par la rupture du canal radial, les chambres flagellées seules sont tapissées de choanocytes. Des chambres flagellées, des canaux ex-courants conduisent dans un spongocoel.
Les canaux incurrents et ex-currents peuvent être complexes et ramifiés. Le spongocoel s’ouvre par un seul osculum. Le parcours du courant d’eau est ostia→ espaces sous-dermiques → canaux incurrents → prosopyles → chambres flagellées → apopyles → canaux ex-currents → spongocoel → osculum → sortie.
Fonctions du système de canaux :
Le système de canaux aide les éponges à se nourrir, respirer, excréter et se reproduire. Le courant d’eau qui circule dans le système de canaux apporte la nourriture et l’oxygène et évacue le dioxyde de carbone, les déchets azotés et les fèces.
Il transporte les spermatozoïdes d’une éponge à l’autre pour la fécondation des ovules. Le système de canaux augmente également la surface des éponges en contact avec l’eau et, ainsi, permet aux éponges d’augmenter leur volume car le rapport surface volume doit rester fixe.
Squelette chez les éponges :
Selon Haeckel peu d’éponges habitant les grands fonds ont un pseudo-squelette composé entièrement de corps étrangers sans aucun élément sécrété par l’éponge elle-même. En revanche, la grande majorité des éponges possèdent un véritable squelette appelé auto squelette composé d’éléments sécrétés par l’éponge elle-même. L’auto-squelette chez les éponges sont soit des spicules, soit de la spongine, soit une combinaison des deux.

Reproduction chez les éponges :
Les éponges se reproduisent à la fois asexuellement et sexuellement.
1. Reproduction asexuée :
La reproduction asexuée se produit chez tous les Porifères. Elle se fait par régénération, formation de corps réducteurs, bourgeonnement et formation de gemmules.
(i) Régénération :
Le pouvoir de régénération est très grand chez les éponges, toute partie coupée régénère l’éponge entière. Si l’éponge est macérée et pressée à travers un tissu de soie fin, ses cellules et grappes de cellules passeront à travers, celles-ci peuvent régénérer de nouvelles éponges. Le pouvoir de régénération est utilisé pour la culture de l’éponge de bain de manière industrielle.
(ii) La formation de corps réducteurs :
Une autre méthode très inhabituelle de reproduction asexuée est la formation de corps de réduction.
De nombreuses éponges d’eau douce et marines se désintègrent dans des circonstances défavorables. L’éponge qui se désintègre s’effondre généralement en laissant de petites boules arrondies, appelées corps de réduction. Chaque corps de réduction consiste en une masse interne d’amibocytes, recouverte extérieurement par un pinacoderme. Lorsque les conditions favorables reviennent, ces corps de réduction se développent en de nouvelles éponges complètes.
(iii) Le bourgeonnement :
Dans les éponges, le bourgeonnement a lieu de diverses manières.
(a) Bourgeonnement exogène :
Une éponge forme des bourgeons externes de manière végétative à la base des branches, formant ainsi, un groupe d’individus. Finalement, les bourgeons se resserrent à partir du parent et chacun forme une nouvelle éponge.
(b) Bourgeonnement endogène :
Des corps reproducteurs asexués appelés gemmules se forment de façon interne chez toutes les éponges d’eau douce et certaines éponges marines.
Les archéocytes se rassemblent en groupes dans le mésogloé, ils deviennent alors multinucléés, ils se remplissent également de protéines comme matière alimentaire de réserve qui est fournie aux archéocytes par des cellules nourricières spéciales, les trophocytes. Certaines amibocytes encerclent cette masse d’archéocytes chargés de nourriture et sécrètent une coquille dure à double couche, la coquille possède une petite sortie ou micropyle.
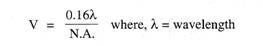
Puis certains scléroblastes sécrètent des spicules qui sont placés radialement entre deux couches de la coquille, certains spicules font saillie à l’extérieur de la coquille, les spicules chez Spongilla sont des spicules monaxons mais chez d’autres ce sont des amphidiscs. Les amphidiscs sont des tiges droites avec des côtés épineux et un anneau de crochets à chaque extrémité.
Ainsi, une gemmule se forme après quoi les amoebocytes, scléroblastes et trophocytes environnants partent. Les gemmules se forment en grand nombre en automne après quoi l’éponge se désagrège, elles restent dans les restes d’éponge ou se libèrent, dans tous les cas elles tombent au fond. Les gemmules peuvent résister à des conditions défavorables, mais elles éclosent lorsque le printemps arrive.
Lors de l’éclosion, les archéocytes ruissellent hors du micropyle, puis ces archéocytes multinucléés se divisent pour former des archéocytes non nucléés et de petites cellules appelées histoblastes.
Les histoblastes par différenciation et réarrangement forment les porocytes de l’épiderme, les choanocytes et le revêtement interne de l’endoderme ; les archéocytes modifiés du scléroblaste qui sécrète des spicules. Environ une semaine après l’éclosion, une jeune éponge entoure la coquille vide de la gemmule. Les archéocytes non nucléés restent embryonnaires.

2. Reproduction sexuée :
Les éponges n’ont pas d’organes sexuels mais les amoebocytes forment des cellules sexuelles dans le mésenchyme ; d’abord les œufs sont produits et plus tard les spermatozoïdes, donc l’éponge est protogyne dans laquelle la fécondation croisée a lieu. Les amoebocytes se remplissent de nourriture et deviennent grands, ils deviennent ronds pour former des œufs.
D’autres amoebocytes se divisent pour produire un grand nombre de spermatozoïdes, un spermatozoïde a une tête ovale et une longue queue effilée.
Certains travailleurs affirment la formation de cellules sexuelles à partir d’archéocytes ou même de choanocytes. Les œufs d’une éponge sont fécondés par les spermatozoïdes d’une autre éponge pour former des zygotes. Lors de la fécondation croisée, le spermatozoïde pénètre probablement dans un choanocyte qui fusionne avec l’ovule, libérant ainsi le spermatozoïde qui fusionne ensuite avec l’ovule. Le zygote sécrète une enveloppe appelée capsule de couvain qui enferme le zygote.

Développement des éponges :
Le zygote subit un clivage holoblastique mais inégal ; dans la division holoblastique, le zygote est complètement segmenté.
Les trois premières divisions sont verticales qui forment huit cellules pyramidales, puis une division horizontale forme huit petites cellules supérieures au pôle animal, et huit grandes cellules inférieures au pôle végétal. Les petites cellules supérieures se divisent rapidement, deviennent claires et acquièrent des flagelles, les cellules inférieures se divisent lentement et deviennent granuleuses.
Ceci forme une blastula qui possède une cavité blastocoel à l’intérieur. La blastula est appelée amphiblastula après la formation des flagelles. Jusqu’ici le développement a lieu dans le corps de l’éponge, maintenant l’amphiblastula trouve son chemin dans les canaux ex-courants et quitte le parent par l’osculum.
L’amphiblastula nage librement pendant quelques heures, puis les cellules flagellées supérieures s’invaginent dans le blastocœle et elles sont repoussées par les cellules granuleuses inférieures ; cela forme une gastrula qui ressemble à une coupe avec une couche extérieure de cellules granuleuses appelée ectoderme, et une couche intérieure de cellules flagellées appelée endoderme, elle possède une grande ouverture, le blastopore.
Cet embryon se fixe à un objet solide par son extrémité blastoporale et il commence à grandir. Les deux couches d’ectoderme et d’endoderme sécrètent le mésogloéa et ses amibocytes. Selon certains, le mésogloéa n’est sécrété que par les choanocytes (endoderme). La paroi s’épaissit et se replie pour former des canaux, les perforations forment des ostia et des osculums.

Les couches germinales des éponges ne sont pas équivalentes à l’ectoderme et à l’endoderme des Métazoaires, car l’ectoderme externe de l’éponge a été formé par les cellules granuleuses inférieures du pôle végétal, et l’endoderme est formé par les cellules flagellées supérieures du pôle animal. Chez les Métazoaires, les cellules du pôle animal deviennent l’ectoderme, et celles du pôle végétal forment l’endoderme.
Nature animale des éponges :
Pendant longtemps, les éponges n’ont pas été considérées comme des animaux, exactement à cause de leur apparence non animale ; elles sont sessiles et ne possèdent pas la capacité d’attraper la nourriture ou de se débarrasser de leurs déchets et ne présentent plus ou moins aucune réponse aux stimuli.
Cependant, c’est Robert Grant (1836) qui a reconnu la nature animale des éponges en suggérant que le courant d’eau circule à travers un chemin défini dans le corps des éponges ; entrant dans le corps de l’éponge par des pores minuscules appelés ostia, dispersés sur toute la surface du corps et sortant par des trous plus grands appelés oscula.
En fait, le mode de nutrition holozoïque des éponges ; elles se nourrissent de particules alimentaires solides venant le long du courant d’eau, dépourvues de paroi cellulaire cellulosique, et la présence de larves ciliées nageant librement dans le cycle de vie ressemblant à ceux d’autres animaux marins a clairement suggéré que les éponges sont des animaux et rien d’autre.
Affinités des éponges :
Il y a eu une grande controverse sur la nature et les affinités des éponges depuis qu’elles ont été découvertes. Aristote (384-322 av. J.-C.) a été le premier à les reconnaître comme des animaux. Mais les biologistes après Aristote les ont cru pendant des siècles comme des plantes en raison de leur habitude sessile et de leur nature insensible.
Ellis (1765) a souligné que le courant d’eau continu entre dans le corps des éponges et en est expulsé ; cela a été confirmé par Robert Grant (1836). Cependant, Linnaeus, Lamarck et Cuvier les plaçaient avec les coelentérés dans Zoophyta. Robert Grant (1836) a finalement établi un groupe séparé Porifera pour les éponges. Ainsi, elles présentent des affinités avec les protozoaires ainsi qu’avec les métazoaires.
Affinités avec les Protozoaires :
Resemblances :
Les éponges présentent des ressemblances avec les Protozoaires coloniaux en ayant les caractéristiques suivantes :
1. Absence de cavité digestive et présence d’une digestion intracellulaire.
2. présence de cellules à collet et de cellules amiboïdes comme celles des flagellés coloniaux.
3. la production du squelette par une seule cellule ou un groupe de cellules.
4. Les cellules du corps de l’éponge sont interdépendantes dans leur fonction.
5. Un processus connu sous le nom d’inversion se produit chez les larves d’amphiblastula comme celles de Volvox. Les larves d’éponges flagellées sont considérées comme le vestige de quelque ancêtre protozoaire colonial flagellé.
Différences :
Les éponges diffèrent des protozoaires en présentant les caractéristiques suivantes :
1. Le système de canaux.
2. le squelette caractéristique.
3. le développement d’un corps multicellulaire à partir d’un œuf fécondé par clivage ; et dans cette caractéristique, les éponges deviennent plus complexes qu’un protozoaire colonial et ressemblent avec les Métazoaires. Par conséquent, son affinité avec les protozoaires ne tient plus.
Affinités avec les Métazoaires :
Resemblances :
Parmi les Métazoaires, les coelentérés présentent certaines similitudes avec les éponges dans les caractéristiques suivantes :
1. Tous deux ont une habitude sédentaire.
2. Tous deux sont diploblastiques et acoelomates. Une couche de mésogloéa non cellulaire se trouve entre l’ectoderme et l’endoderme. Le mésoderme est absent dans les deux cas.
3. Le spongocoel des éponges s’ouvrant par l’osculum terminal peut être comparé à la cavité gastro-vasculaire, s’ouvrant sur l’extérieur par l’hypos tome terminal des cœlentérés.
4 La reproduction asexuée se produit dans les deux cas et les colonies se forment par bourgeonnement.
5. La larve parenchymateuse des éponges est comparable à la planula des coelentérés.
Différences :
Les spongiaires diffèrent des Métazoaires en présentant les caractéristiques suivantes :
1. Chez les spongiaires, les cellules sont comparativement moins spécialisées et interdépendantes que les cellules des Métazoaires.
2. Les spongiaires possèdent des choanocytes qui ne se trouvent chez aucun Métazoaire.
3. Les éponges sont au niveau de la construction cellulaire mais les Métazoaires sont au niveau de la construction des tissus et des organes.
4. Les éponges sont diploblastiques, alors que les Métazoaires (sauf les Coelentérés) sont triploblastiques.
5. Les éponges ne possèdent pas d’extrémité antérieure ou de tête comme celles des Métazoaires. L’osculum joue uniquement le rôle d’ouverture exhalante.
5. Le corps des éponges est perforé par de nombreux pores minuscules et elles possèdent un système unique de canaux dans leur corps ; tout cela ne se trouve jamais chez les Métazoaires.
6. Les éponges n’ont pas de tissus nerveux pour coordonner les activités du corps.
7. Il y a une absence totale de cavité digestive chez les éponges, contrairement à celles des Métazoaires.
8. Les éponges ont un plus grand nombre de graisses et d’acides gras qui sont d’un poids moléculaire plus élevé que ceux trouvés dans les Métazoaires.
9. Dans le développement des spongiaires, les cellules supérieures flagellées de l’embryon, situées au pôle animal, deviennent l’endoderme et les cellules granuleuses du pôle végétal deviennent l’ectoderme ; chez aucun Métazoaire une telle inversion ne se produit au cours du développement.
10. En raison de ces différences, les éponges ne peuvent pas être regroupées avec les Métazoaires et, par conséquent, un phylum distinct a été créé pour les éponges.
Ainsi, en dépit de ses ressemblances avec les Protozoaires et les Métazoaires, on pense que les éponges ont divergé de la ligne principale de l’évolution des Métazoaires et qu’elles constituent un phylum sans issue. Considérant cela, un sous-royaume distinct Parazoa a été créé pour inclure les éponges et, par conséquent, elles ont une position phylogénétique isolée.
Importance économique des éponges :
Les éponges ont une grande importance économique, à la fois bénéfique et nuisible, pour l’humanité.
A. Les éponges bénéfiques :
Les éponges sont bénéfiques pour l’homme et les autres animaux de la manière suivante :
1. Comme nourriture :
Les éponges sont rarement prises comme nourriture par les autres animaux à cause de leur mauvais goût, de leur odeur et de leurs spicules pointus. Cependant, on trouve des crustacés menant une vie parasitaire sur elles et certains mollusques comme les nudibranches en dépendent pour leur alimentation.
2. en tant que commensaux :
Les éponges servent de maisons protectrices pour plusieurs animaux comme les crustacés, les vers, les mollusques, les petits poissons, etc. car leurs ennemis ne peuvent pas se nourrir des éponges. En plus de la protection, les animaux vivant à l’intérieur du corps de l’éponge obtiennent une riche alimentation à partir de l’eau qui circule à travers eux. Ces animaux ne causent aucun dommage apparent aux éponges, ils sont considérés comme des commensaux.
3. autres utilisations :
Le squelette séché et fibreux de nombreuses éponges comme Spongia, Hippospongia et Euspongia est utilisé pour le bain, le polissage, le lavage des voitures, des murs, des meubles et le récurage des sols, etc. Le squelette de certaines éponges comme Euplectella ont une grande valeur commerciale et sont utilisées comme pièces décoratives.
B. Les éponges nuisibles :
Seulement quelques éponges sont nocives. Elles peuvent provoquer la mort de certains animaux sessiles en se développant sur eux et en coupant leur alimentation et leur oxygène. Les éponges foreuses, comme Cliona se fixent sur les coquilles des huîtres, des palourdes et des bernacles, etc. Elle fore dans les coquilles de ces animaux et les détruit complètement.
Les éponges foreuses causent également de grands dommages aux bancs d’huîtres. Les éponges foreuses détruisent aussi les rochers en y pénétrant et en les brisant en morceaux.
